L'ADN est brièvement porteur d'informations héréditaires. A1
Informations génétiques dans une cellule. Gènes, code génétique et ses propriétés. Nature matricielle des réactions de biosynthèse. Biosynthèse des protéines et des acides nucléiques
Informations génétiques dans une cellule
La reproduction des êtres humains est l'une des propriétés fondamentales des êtres vivants. Grâce à ce phénomène, il existe des similitudes non seulement entre les organismes, mais aussi entre les cellules individuelles, ainsi qu'entre leurs organites (mitochondries et plastes). La base matérielle de cette similitude est le transfert d'informations génétiques cryptées dans la séquence nucléotidique de l'ADN, qui s'effectue via les processus de réplication de l'ADN (auto-duplication). Toutes les caractéristiques et propriétés des cellules et des organismes sont réalisées grâce à des protéines dont la structure est principalement déterminée par la séquence de nucléotides d'ADN. La biosynthèse joue donc un rôle primordial dans les processus métaboliques. acides nucléiques et l'écureuil. L'unité structurelle de l'information héréditaire est le gène.
Gènes, code génétique et ses propriétés
Les informations héréditaires dans une cellule ne sont pas monolithiques, elles sont divisées en « mots » distincts : les gènes.
Gène est une unité élémentaire d’information génétique.
Les travaux sur le programme « Génome humain », menés simultanément dans plusieurs pays et achevés au début de ce siècle, nous ont permis de comprendre qu'une personne ne possède qu'environ 25 à 30 000 gènes, mais que les informations provenant de la plupart de notre ADN n'est jamais lu, car il contient grande quantité des sections dénuées de sens, des répétitions et des gènes codant pour des traits qui ont perdu leur sens pour les humains (queue, poils, etc.). En outre, un certain nombre de gènes responsables du développement de maladies héréditaires, ainsi que des gènes cibles de médicaments, ont été décryptés. Cependant utilisation pratique Les résultats obtenus lors de la mise en œuvre de ce programme sont reportés jusqu'à ce que les génomes d'un plus grand nombre de personnes soient déchiffrés et qu'il devienne clair en quoi ils diffèrent.
Les gènes qui codent pour la structure primaire d'une protéine, d'un ribosomal ou d'un ARN de transfert sont appelés de construction, et les gènes qui assurent l'activation ou la suppression des informations de lecture provenant de gènes structurels - réglementaire. Cependant, même les gènes structurels contiennent des régions régulatrices.
Les informations héréditaires des organismes sont cryptées dans l'ADN sous la forme de certaines combinaisons de nucléotides et de leur séquence - code génétique. Ses propriétés sont : la tripleté, la spécificité, l'universalité, la redondance et la non-recoupement. De plus, il n’y a aucun signe de ponctuation dans le code génétique.
Chaque acide aminé est codé dans l'ADN par trois nucléotides - triolet, par exemple, la méthionine est codée par le triplet TAC, c'est-à-dire que le code est un triplet. En revanche, chaque triplet ne code qu'un seul acide aminé, ce qui constitue sa spécificité ou son absence d'ambiguïté. Code génétique est universel pour tous les organismes vivants, c'est-à-dire que les informations héréditaires sur les protéines humaines peuvent être lues par les bactéries et vice versa. Cela indique l'unité d'origine monde organique. Cependant, 64 combinaisons de trois nucléotides correspondent à seulement 20 acides aminés, de sorte qu'un acide aminé peut être codé par 2 à 6 triplets, c'est-à-dire que le code génétique est redondant ou dégénéré. Trois triplés n'ont pas d'acides aminés correspondants, ils sont appelés arrêter les codons, puisqu'ils indiquent la fin de la synthèse de la chaîne polypeptidique.
La séquence de bases dans les triplets d'ADN et les acides aminés qu'ils codent
*Codon Stop, indiquant la fin de la synthèse de la chaîne polypeptidique.
Abréviations des noms d’acides aminés :
Ala - alanine
Arg - arginine
Asn - asparagine
Asp - acide aspartique
Val - valine
Son - histidine
Gly-glycine
Gln-glutamine
Glu - acide glutamique
Île-isoleucine
Leu - leucine
Liz - lysine
Méth - méthionine
Pro-proline
Ser - sérine
Tyr - tyrosine
Tre - thréonine
Tri-tryptophane
Fen - phénylalanine
Cis - cystéine
Si vous commencez à lire des informations génétiques non pas à partir du premier nucléotide du triplet, mais à partir du second, non seulement le cadre de lecture changera - la protéine ainsi synthétisée sera complètement différente non seulement dans la séquence nucléotidique, mais également dans sa structure. et propriétés. Il n'y a pas de signes de ponctuation entre les triplets, il n'y a donc aucun obstacle au déplacement du cadre de lecture, ce qui ouvre un espace pour l'apparition et le maintien des mutations.
Nature matricielle des réactions de biosynthèse
Les cellules bactériennes sont capables de doubler toutes les 20 à 30 minutes, et les cellules eucaryotes, chaque jour et même plus souvent, ce qui nécessite une réplication rapide et précise de l'ADN. De plus, chaque cellule contient des centaines et des milliers de copies de nombreuses protéines, notamment des enzymes, par conséquent, la méthode « fragmentaire » de leur production est inacceptable pour leur reproduction. Une méthode plus progressive est l'estampage, qui permet d'obtenir de nombreuses copies exactes du produit et également de réduire son coût. Pour l'estampage, vous avez besoin d'une matrice à partir de laquelle l'impression est réalisée.
Dans les cellules, le principe de la synthèse de matrices est que de nouvelles molécules de protéines et d'acides nucléiques sont synthétisées conformément au programme intégré dans la structure des molécules préexistantes des mêmes acides nucléiques (ADN ou ARN).
Biosynthèse des protéines et des acides nucléiques
Réplication de l'ADN. L'ADN est un biopolymère double brin dont les monomères sont des nucléotides. Si la biosynthèse de l'ADN se produisait selon le principe de la photocopie, de nombreuses distorsions et erreurs dans les informations héréditaires se produiraient inévitablement, ce qui conduirait finalement à la mort de nouveaux organismes. Par conséquent, le processus de doublement de l’ADN se déroule différemment, de manière semi-conservatrice: la molécule d'ADN se déroule, et une nouvelle chaîne est synthétisée sur chacune des chaînes selon le principe de complémentarité. Le processus d'auto-reproduction d'une molécule d'ADN, assurant une copie précise des informations héréditaires et sa transmission de génération en génération, est appelé réplication(de lat. réplication- répétition). À la suite de la réplication, deux copies absolument exactes de la molécule d'ADN mère sont formées, chacune portant une copie de la molécule d'ADN mère.

Le processus de réplication est en réalité extrêmement complexe, car un certain nombre de protéines y sont impliquées. Certains d'entre eux déroulent la double hélice de l'ADN, d'autres rompent les liaisons hydrogène entre les nucléotides des chaînes complémentaires, d'autres (par exemple, l'enzyme ADN polymérase) sélectionnent de nouveaux nucléotides sur la base du principe de complémentarité, etc. Deux molécules d'ADN formées comme un Le résultat de la réplication diverge en deux lors de la division des cellules filles nouvellement formées.
Les erreurs dans le processus de réplication se produisent extrêmement rarement, mais si elles se produisent, elles sont très rapidement éliminées à la fois par les ADN polymérases et par les enzymes de réparation spéciales, car toute erreur dans la séquence nucléotidique peut entraîner une modification irréversible de la structure et des fonctions de la protéine. et, finalement, nuire à la viabilité d’une nouvelle cellule ou même d’un individu.
Biosynthèse des protéines. Comme le disait figurativement l'éminent philosophe du XIXe siècle F. Engels : « La vie est une forme d'existence de corps protéiques ». La structure et les propriétés des molécules protéiques sont déterminées par leur structure primaire, c'est-à-dire la séquence d'acides aminés codée dans l'ADN. Non seulement l'existence du polypeptide lui-même, mais également le fonctionnement de la cellule dans son ensemble dépendent de l'exactitude de la reproduction de ces informations, c'est pourquoi le processus de synthèse des protéines est d'une grande importance. Il semble que ce soit le processus de synthèse le plus complexe de la cellule, puisqu’il implique jusqu’à trois cents enzymes et autres macromolécules différentes. De plus, cela se produit avec grande vitesse, ce qui nécessite encore plus de précision.
Il y a deux étapes principales dans la biosynthèse des protéines : la transcription et la traduction.
Transcription(de lat. transcription- réécriture) est la biosynthèse de molécules d'ARNm sur une matrice d'ADN.

Étant donné que la molécule d'ADN contient deux chaînes antiparallèles, la lecture des informations des deux chaînes conduirait à la formation d'ARNm complètement différents, leur biosynthèse n'est donc possible que sur l'une des chaînes, qui est dite codante, ou codogène, contrairement à la seconde, non codant ou non codogène. Le processus de réécriture est assuré par une enzyme particulière, l'ARN polymérase, qui sélectionne les nucléotides d'ARN selon le principe de complémentarité. Ce processus peut se produire à la fois dans le noyau et dans les organites qui possèdent leur propre ADN - les mitochondries et les plastes.
Les molécules d'ARNm synthétisées lors de la transcription subissent un processus complexe de préparation à la traduction (les ARNm mitochondriaux et plastidiens peuvent rester à l'intérieur des organites, où se produit la deuxième étape de la biosynthèse des protéines). Au cours du processus de maturation de l'ARNm, les trois premiers nucléotides (AUG) et une queue de nucléotides adényliques y sont attachés, dont la longueur détermine le nombre de copies de la protéine pouvant être synthétisées sur une molécule donnée. Ce n’est qu’alors que les ARNm matures quittent le noyau par les pores nucléaires.
En parallèle, le processus d'activation des acides aminés se produit dans le cytoplasme, au cours duquel l'acide aminé rejoint l'ARNt libre correspondant. Ce processus est catalysé par une enzyme spéciale et nécessite de l'ATP.
Diffuser(de lat. diffuser- transfert) est la biosynthèse d'une chaîne polypeptidique sur une matrice d'ARNm, au cours de laquelle l'information génétique est traduite en séquence d'acides aminés de la chaîne polypeptidique.

La deuxième étape de la synthèse protéique se produit le plus souvent dans le cytoplasme, par exemple sur le RE rugueux. Pour son apparition, la présence de ribosomes, l'activation des ARNt, au cours de laquelle ils fixent les acides aminés correspondants, la présence d'ions Mg2+, ainsi que des conditions environnementales optimales (température, pH, pression, etc.) sont nécessaires.
Pour démarrer la diffusion ( initiation) une petite sous-unité ribosomale est attachée à une molécule d'ARNm prête à être synthétisée, puis, selon le principe de complémentarité avec le premier codon (AUG), un ARNt portant l'acide aminé méthionine est sélectionné. Ce n’est qu’après que la grande sous-unité ribosomale s’attache. Dans le ribosome assemblé, il y a deux codons d'ARNm, dont le premier est déjà occupé. Un deuxième ARNt, portant également un acide aminé, est ajouté au codon qui lui est adjacent, après quoi une liaison peptidique est formée entre les résidus d'acides aminés à l'aide d'enzymes. Le ribosome déplace un codon de l'ARNm ; le premier ARNt libéré d'un acide aminé retourne dans le cytoplasme après l'acide aminé suivant, et un fragment de la future chaîne polypeptidique est suspendu, pour ainsi dire, à l'ARNt restant. L'ARNt suivant est attaché au nouveau codon qui se trouve dans le ribosome, le processus est répété et étape par étape la chaîne polypeptidique s'allonge, c'est-à-dire élongation.
Fin de la synthèse protéique ( Résiliation) se produit dès qu’une séquence nucléotidique spécifique est rencontrée dans la molécule d’ARNm qui ne code pas pour un acide aminé (codon stop). Après cela, le ribosome, l'ARNm et la chaîne polypeptidique sont séparés et la protéine nouvellement synthétisée acquiert la structure appropriée et est transportée vers la partie de la cellule où elle remplira ses fonctions.
La traduction est un processus très gourmand en énergie, puisque l'énergie d'une molécule d'ATP est consommée pour attacher un acide aminé à l'ARNt, et plusieurs autres sont utilisées pour déplacer le ribosome le long de la molécule d'ARNm.
Pour accélérer la synthèse de certaines molécules protéiques, plusieurs ribosomes peuvent être successivement attachés à une molécule d'ARNm, qui forment une structure unique - polysome.
Une cellule est l'unité génétique d'un être vivant. Les chromosomes, leur structure (forme et taille) et leurs fonctions. Le nombre de chromosomes et la constance de leur espèce. Cellules somatiques et germinales. Cycle de vie cellulaire : interphase et mitose. La mitose est la division des cellules somatiques. Méiose. Phases de mitose et de méiose. Développement de cellules germinales chez les plantes et les animaux. La division cellulaire est la base de la croissance, du développement et de la reproduction des organismes. Le rôle de la méiose et de la mitose
Cellule - unité génétique des êtres vivants
Malgré le fait que les acides nucléiques soient porteurs d'informations génétiques, la mise en œuvre de ces informations est impossible en dehors de la cellule, ce qui est facilement prouvé par l'exemple des virus. Ces organismes, qui ne contiennent souvent que de l'ADN ou de l'ARN, ne peuvent pas se reproduire de manière indépendante ; pour ce faire, ils doivent utiliser l'appareil héréditaire de la cellule. Ils ne peuvent même pas pénétrer dans une cellule sans l’aide de la cellule elle-même, sauf par l’utilisation de mécanismes de transport membranaire ou en raison de dommages cellulaires. La plupart des virus sont instables : ils meurent après seulement quelques heures d’exposition à l’air libre. Par conséquent, une cellule est une unité génétique d’un être vivant, possédant ensemble minimum les éléments de conservation, de modification et de mise en œuvre de l'information héréditaire, ainsi que sa transmission à la descendance.
La plupart des informations génétiques d'une cellule eucaryote se trouvent dans le noyau. La particularité de son organisation est que, contrairement à l'ADN d'une cellule procaryote, les molécules d'ADN des eucaryotes ne sont pas fermées et forment des complexes complexes avec des protéines - les chromosomes.
Les chromosomes, leur structure (forme et taille) et leurs fonctions
Chromosome(du grec chrome- couleur, coloration et soma- corps) est la structure du noyau cellulaire, qui contient des gènes et porte certaines informations héréditaires sur les caractéristiques et les propriétés de l'organisme.
Parfois, les molécules d’ADN circulaires des procaryotes sont également appelées chromosomes. Les chromosomes sont capables de s'autodupliquer ; ils ont une individualité structurelle et fonctionnelle et la conservent au fil des générations. Chaque cellule transporte toutes les informations héréditaires du corps, mais seule une petite partie y travaille.
La base d'un chromosome est une molécule d'ADN double brin remplie de protéines. Chez les eucaryotes, les protéines histones et non histones interagissent avec l'ADN, alors que chez les procaryotes, les protéines histones sont absentes.
Les chromosomes sont mieux visibles au microscope optique lors de la division cellulaire, lorsque, à la suite du compactage, ils prennent l'apparence de corps en forme de bâtonnets séparés par un étranglement primaire - centromère - sur les épaules. Sur un chromosome, il peut aussi y avoir constriction secondaire, qui dans certains cas sépare ce qu'on appelle Satellite. Les extrémités des chromosomes sont appelées télomères. Les télomères empêchent les extrémités des chromosomes de se coller et assurent leur fixation à la membrane nucléaire dans une cellule qui ne se divise pas. Au début de la division, les chromosomes sont doublés et se composent de deux chromosomes filles - chromatide, fixé au centromère.

Selon leur forme, les chromosomes sont divisés en chromosomes à bras égaux, à bras inégaux et en forme de bâtonnet. Les tailles des chromosomes varient considérablement, mais le chromosome moyen a des dimensions de 5 $×$ 1,4 microns.
Dans certains cas, les chromosomes, suite à de nombreuses duplications d'ADN, contiennent des centaines et des milliers de chromatides : ces chromosomes géants sont appelés polytène. On les trouve dans les glandes salivaires des larves de drosophile, ainsi que dans les glandes digestives des vers ronds.
Le nombre de chromosomes et la constance de leur espèce. Cellules somatiques et germinales
Selon la théorie cellulaire, une cellule est une unité de structure, d’activité vitale et de développement d’un organisme. Ainsi, des fonctions aussi importantes des êtres vivants que la croissance, la reproduction et le développement de l'organisme sont assurées au niveau cellulaire. Cellules Organismes multicellulaires peut être divisé en somatique et sexuel.
Cellules somatiques- ce sont toutes les cellules du corps formées à la suite d'une division mitotique.
L'étude des chromosomes a permis d'établir que les cellules somatiques de l'organisme de chaque espèce biologique sont caractérisées par un nombre constant de chromosomes. Par exemple, une personne en possède 46. L’ensemble des chromosomes des cellules somatiques est appelé diploïde(2n), ou double.

Cellules sexuelles, ou gamètes, sont des cellules spécialisées utilisées pour la reproduction sexuée.
Les gamètes contiennent toujours la moitié moins de chromosomes que les cellules somatiques (chez l'homme - 23), c'est pourquoi l'ensemble des chromosomes des cellules germinales est appelé haploïde(n), ou célibataire. Sa formation est associée à la division cellulaire méiotique.
La quantité d'ADN dans les cellules somatiques est désignée par 2c et dans les cellules sexuelles - 1c. La formule génétique des cellules somatiques s'écrit 2n2c et celle des cellules sexuelles - 1n1c.
Dans les noyaux de certaines cellules somatiques, le nombre de chromosomes peut différer de leur nombre dans les cellules somatiques. Si cette différence est supérieure à un, deux, trois, etc. ensembles haploïdes, alors ces cellules sont appelées polyploïde(tri-, tétra-, pentaploïde, respectivement). Dans ces cellules, les processus métaboliques se déroulent généralement de manière très intensive.
Le nombre de chromosomes en lui-même n'est pas une caractéristique spécifique à l'espèce, puisque différents organismes peuvent avoir un nombre égal de chromosomes, mais que des organismes apparentés peuvent en avoir un nombre différent. Par exemple, le plasmodium du paludisme et l'ascaris du cheval ont chacun deux chromosomes, tandis que les humains et les chimpanzés en ont respectivement 46 et 48.
Les chromosomes humains sont divisés en deux groupes : les autosomes et les chromosomes sexuels (hétérochromosomes). Autosome dans les cellules somatiques humaines, il y a 22 paires, elles sont les mêmes pour les hommes et les femmes, et chromosomes sexuels un seul couple, mais c'est lui qui détermine le sexe de l'individu. Il existe deux types de chromosomes sexuels - X et Y. Les cellules du corps des femmes portent deux chromosomes X et celles des hommes - X et Y.
Caryotype- il s'agit d'un ensemble de caractéristiques de l'ensemble chromosomique d'un organisme (le nombre de chromosomes, leur forme et leur taille).
L'enregistrement conditionnel d'un caryotype comprend le nombre total de chromosomes, les chromosomes sexuels et les écarts possibles dans l'ensemble des chromosomes. Par exemple, le caryotype d'un homme normal s'écrit 46,XY, et le caryotype femme normale- 46, XX.
Cycle de vie cellulaire : interphase et mitose
Les cellules ne se reproduisent pas à chaque fois, elles se forment uniquement à la suite de la division des cellules mères. Après la division, les cellules filles ont besoin d'un certain temps pour former des organites et acquérir la structure appropriée qui assurerait l'exécution d'une fonction spécifique. Cette période de temps est appelée maturation.
La période de temps allant de l'apparition d'une cellule à la suite d'une division jusqu'à sa division ou sa mort est appelée cycle de vie d'une cellule.
Dans les cellules eucaryotes, le cycle de vie est divisé en deux étapes principales : l’interphase et la mitose.
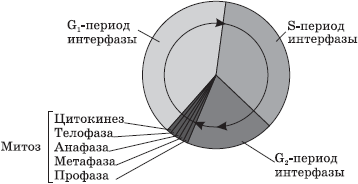
Interphase- il s'agit d'une période du cycle de vie pendant laquelle la cellule ne se divise pas et fonctionne normalement. L'interphase est divisée en trois périodes : périodes G 1 -, S- et G 2 -.
G 1 -période(présynthétique, postmitotique) est une période de croissance et de développement cellulaire au cours de laquelle se produit la synthèse active d'ARN, de protéines et d'autres substances nécessaires au maintien complet de la vie de la cellule nouvellement formée. Vers la fin de cette période, la cellule peut commencer à se préparer à dupliquer son ADN.
DANS Période S(synthétique) le processus de réplication de l'ADN lui-même se produit. La seule partie du chromosome qui ne subit pas de réplication est le centromère, de sorte que les molécules d'ADN résultantes ne divergent pas complètement, mais y restent maintenues ensemble et, au début de la division, le chromosome a une apparence en forme de X. La formule génétique d'une cellule après doublement de l'ADN est 2n4c. Également pendant la période S, les centrioles du centre cellulaire sont doublés.
G 2 -période(postsynthétique, prémitotique) se caractérise par une synthèse intensive d'ARN, de protéines et d'ATP nécessaires au processus de division cellulaire, ainsi que par la séparation des centrioles, des mitochondries et des plastes. Jusqu'à la fin de l'interphase, la chromatine et le nucléole restent clairement distinguables, l'intégrité de l'enveloppe nucléaire n'est pas perturbée et les organites ne changent pas.
Certaines cellules de l'organisme sont capables de remplir leurs fonctions tout au long de la vie de l'organisme (neurones de notre cerveau, cellules musculaires du cœur), tandis que d'autres existent pendant une courte période, après quoi elles meurent (cellules épithéliales intestinales, cellules épidermiques de la peau). Par conséquent, le corps doit constamment subir des processus de division cellulaire et la formation de nouvelles cellules qui remplaceront les cellules mortes. Les cellules capables de se diviser sont appelées tige. Dans le corps humain, on les trouve dans la moelle osseuse rouge, dans les couches profondes de l'épiderme de la peau et à d'autres endroits. En utilisant ces cellules, vous pouvez développer un nouvel organe, obtenir un rajeunissement et également cloner le corps. Les perspectives d'utilisation des cellules souches sont tout à fait claires, mais les aspects moraux et éthiques de ce problème sont toujours en discussion, puisque dans la plupart des cas, on utilise des cellules souches embryonnaires obtenues à partir d'embryons humains tués lors d'un avortement.
La durée de l'interphase dans les cellules végétales et animales est en moyenne de 10 à 20 heures, tandis que la mitose prend environ 1 à 2 heures.
Au cours des divisions successives des organismes multicellulaires, les cellules filles se diversifient de plus en plus à mesure qu'elles lisent les informations provenant de tous. plus gènes.
Certaines cellules cessent de se diviser avec le temps et meurent, ce qui peut être dû à l'achèvement de certaines fonctions, comme dans le cas des cellules épidermiques de la peau et des cellules sanguines, ou à des dommages causés à ces cellules par des facteurs. environnement, en particulier les agents pathogènes. La mort cellulaire génétiquement programmée est appelée apoptose, alors que la mort accidentelle - nécrose.
La mitose est la division des cellules somatiques. Phases de la mitose
Mitose- une méthode de division indirecte des cellules somatiques.
Au cours de la mitose, la cellule passe par une série de phases successives, à la suite desquelles chaque cellule fille reçoit le même ensemble de chromosomes que la cellule mère.
La mitose est divisée en quatre phases principales : prophase, métaphase, anaphase et télophase. Prophase- l'étape la plus longue de la mitose, au cours de laquelle la chromatine se condense, ce qui donne naissance à des chromosomes en forme de X constitués de deux chromatides (chromosomes filles) devenant visibles. Dans ce cas, le nucléole disparaît, les centrioles divergent vers les pôles de la cellule et un fuseau d'achromatine (fuseau de division) à partir de microtubules commence à se former. A la fin de la prophase, la membrane nucléaire se désintègre en vésicules distinctes.

DANS métaphase Les chromosomes sont alignés le long de l'équateur de la cellule avec leurs centromères, auxquels sont attachés les microtubules du fuseau entièrement formé. A ce stade de division, les chromosomes sont les plus compactés et ont une forme caractéristique, ce qui permet d'étudier le caryotype.
DANS anaphase La réplication rapide de l'ADN se produit au niveau des centromères, à la suite de laquelle les chromosomes sont divisés et les chromatides divergent vers les pôles de la cellule, étirés par les microtubules. La répartition des chromatides doit être absolument égale, puisque c'est ce processus qui assure le maintien d'un nombre constant de chromosomes dans les cellules du corps.
Sur la scène télophases les chromosomes filles se rassemblent aux pôles, des membranes déspirales et nucléaires se forment autour d'eux à partir de vésicules et des nucléoles apparaissent dans les noyaux nouvellement formés.
Après la division nucléaire, la division cytoplasmique se produit - cytokinèse, au cours de laquelle se produit une répartition plus ou moins uniforme de tous les organites de la cellule mère.
Ainsi, à la suite de la mitose, deux cellules filles sont formées à partir d'une cellule mère, chacune étant une copie génétique de la cellule mère (2n2c).
Dans les cellules malades, endommagées et vieillissantes et dans les tissus spécialisés du corps, un processus de division légèrement différent peut se produire : l'amitose. Amitose appelée division directe des cellules eucaryotes, dans laquelle la formation de cellules génétiquement équivalentes ne se produit pas, car les composants cellulaires sont inégalement répartis. On le trouve dans les plantes dans l'endosperme et chez les animaux - dans le foie, le cartilage et la cornée de l'œil.
Méiose. Phases de la méiose
Méiose est une méthode de division indirecte des cellules germinales primaires (2n2c), qui aboutit à la formation de cellules haploïdes (1n1c), le plus souvent des cellules germinales.
Contrairement à la mitose, la méiose est constituée de deux divisions cellulaires successives, chacune étant précédée d'une interphase. La première division de la méiose (méiose I) est appelée réductionniste, puisque dans ce cas le nombre de chromosomes est réduit de moitié, et la deuxième division (méiose II) - équationnel, puisque dans son processus le nombre de chromosomes est préservé.

Interphase I se déroule comme une interphase de mitose. Méiose I est divisé en quatre phases : prophase I, métaphase I, anaphase I et télophase I. B prophase I Deux processus importants se produisent : la conjugaison et le croisement. Conjugaison- Il s'agit du processus de fusion de chromosomes homologues (appariés) sur toute la longueur. Les paires de chromosomes formées lors de la conjugaison sont conservées jusqu'à la fin de la métaphase I.
Traverser- échange mutuel de régions homologues de chromosomes homologues. À la suite du croisement, les chromosomes reçus par le corps des deux parents acquièrent de nouvelles combinaisons de gènes, ce qui provoque l'apparition d'une progéniture génétiquement diversifiée. A la fin de la prophase I, comme lors de la prophase de mitose, le nucléole disparaît, les centrioles divergent vers les pôles de la cellule et la membrane nucléaire se désintègre.

DANS métaphase I des paires de chromosomes s'alignent le long de l'équateur de la cellule et des microtubules fusiformes sont attachés à leurs centromères.
DANS anaphase I Les chromosomes homologues entiers, constitués de deux chromatides, divergent vers les pôles.
DANS télophase I Les membranes nucléaires se forment autour des amas de chromosomes aux pôles de la cellule et des nucléoles se forment.
Cytocinèse I assure la séparation des cytoplasmes des cellules filles.
Les cellules filles (1n2c) formées à la suite de la méiose I sont génétiquement hétérogènes, puisque leurs chromosomes, dispersés de manière aléatoire vers les pôles cellulaires, contiennent des gènes différents.
Caractéristiques comparatives de la mitose et de la méiose
| Signe | Mitose | Méiose | |
| Quelles cellules commencent à se diviser ? | Somatique (2n) | Cellules germinales primaires (2n) | |
| Nombre de divisions | 1 | 2 | |
| Combien et quel type de cellules se forment lors de la division ? | 2 somatique (2n) | 4 sexuel (n) | |
| Interphase | Préparer la cellule à la division, doubler l'ADN | Très court, le doublement de l'ADN ne se produit pas | |
| Phases | Méiose I | Méiose II | |
| Prophase | Une condensation chromosomique, une disparition du nucléole, une désintégration de la membrane nucléaire, une conjugaison et un croisement peuvent survenir. | Condensation chromosomique, disparition du nucléole, désintégration de la membrane nucléaire | |
| Métaphase | Des paires de chromosomes sont situées le long de l'équateur, un fuseau se forme | Les chromosomes s'alignent le long de l'équateur, un fuseau se forme | |
| Anaphase | Les chromosomes homologues de deux chromatides se déplacent vers les pôles | Les chromatides se dirigent vers les pôles | |
| Télophase | Les chromosomes déspirent, de nouvelles membranes nucléaires et de nouveaux nucléoles se forment | Les chromosomes déspirent, de nouvelles membranes nucléaires et de nouveaux nucléoles se forment | |
Interphase II très court, car le doublement de l'ADN ne s'y produit pas, c'est-à-dire qu'il n'y a pas de période S.
Méiose IIégalement divisé en quatre phases : prophase II, métaphase II, anaphase II et télophase II. DANS prophase II les mêmes processus se produisent que dans la prophase I, à l'exception de la conjugaison et du croisement.
DANS métaphase II les chromosomes sont situés le long de l’équateur de la cellule.
DANS anaphase II les chromosomes sont divisés au niveau des centromères et les chromatides sont étirées vers les pôles.
DANS télophase II Les membranes nucléaires et les nucléoles se forment autour d’amas de chromosomes filles.
Après cytokinèse II La formule génétique des quatre cellules filles est 1n1c, mais elles possèdent toutes un ensemble de gènes différent, résultat du croisement et de la combinaison aléatoire de chromosomes des organismes maternels et paternels dans les cellules filles.
Développement des cellules germinales chez les plantes et les animaux
Gamétogenèse(du grec gamète- épouse, gamètes- mon mari et genèse- origine, émergence) est le processus de formation de cellules germinales matures.
Puisque la reproduction sexuée nécessite le plus souvent deux individus - une femelle et un mâle, produisant des cellules sexuelles différentes - des ovules et des spermatozoïdes, les processus de formation de ces gamètes doivent être différents.
La nature du processus dépend dans une large mesure du fait qu'il se déroule en usine ou cellule animale, puisque chez les plantes, lors de la formation des gamètes, seule la mitose se produit, et chez les animaux, la mitose et la méiose se produisent à la fois.
Développement des cellules germinales chez les plantes. Chez les angiospermes, la formation des gamètes mâles et femelles se produit dans diverses pièces fleur - respectivement étamines et pistils.
Avant la formation des cellules reproductrices mâles - microgamétogenèse(du grec micros- petit) - arrive microsporogenèse, c'est-à-dire la formation de microspores dans les anthères des étamines. Ce processus est associé à la division méiotique de la cellule mère, qui donne naissance à quatre microspores haploïdes. La microgamétogenèse est associée à la division mitotique de la microspore, produisant un gamétophyte mâle composé de deux cellules - un grand végétatif(siphonogène) et peu profond génératif. Après division, le gamétophyte mâle se recouvre de membranes denses et forme un grain de pollen. Dans certains cas, même pendant le processus de maturation du pollen, et parfois seulement après transfert sur le stigmate du pistil, la cellule générative se divise par mitose pour former deux cellules germinales mâles immobiles - sperme. Après la pollinisation, un tube pollinique se forme à partir de la cellule végétative, à travers lequel les spermatozoïdes pénètrent dans l'ovaire du pistil pour la fécondation.

Le développement des cellules germinales femelles chez les plantes est appelé mégagamétogenèse(du grec mégas- grand). Il se produit dans l'ovaire du pistil, qui est précédé par mégasporogenèse, à la suite de quoi quatre mégaspores sont formées à partir de la cellule mère de la mégaspore située dans le nucelle par division méiotique. L'une des mégaspores se divise mitotiquement trois fois, donnant le gamétophyte femelle - un sac embryonnaire à huit noyaux. Avec la séparation ultérieure des cytoplasmes des cellules filles, l'une des cellules résultantes devient un œuf, sur les côtés duquel se trouvent ce qu'on appelle les synergides, à l'extrémité opposée du sac embryonnaire se forment trois antipodes et au centre , à la suite de la fusion de deux noyaux haploïdes, une cellule centrale diploïde se forme.

Développement de cellules germinales chez les animaux. Chez les animaux, il existe deux processus de formation de cellules germinales : la spermatogenèse et l'ovogenèse.
Spermatogenèse(du grec sperme, spermatozoïdes- des graines et genèse- origine, apparition) est le processus de formation de cellules germinales mâles matures - le sperme. Chez l'homme, elle se produit dans les testicules, ou testicules, et se divise en quatre périodes : reproduction, croissance, maturation et formation.

DANS saison des amours Les cellules germinales primordiales se divisent par mitose, entraînant la formation de cellules diploïdes. spermatogonie. DANS période de croissance les spermatogonies accumulent des nutriments dans le cytoplasme, augmentent de taille et se transforment en spermatocytes primaires, ou Spermatocytes de 1er ordre. Ce n'est qu'après cela qu'ils entrent en méiose ( période de maturation), à la suite de quoi les deux premiers sont formés spermatocyte secondaire, ou spermatocyte de 2ème ordre, puis - quatre cellules haploïdes avec encore une quantité assez importante de cytoplasme - spermatides. DANS période de formation ils perdent presque tout leur cytoplasme et forment un flagelle, se transformant en spermatozoïdes.
Sperme, ou animés, - de très petites cellules reproductrices mâles mobiles dotées d'une tête, d'un cou et d'une queue.

DANS tête, en plus du noyau, est acrosome- un complexe de Golgi modifié, qui assure la dissolution des membranes de l'œuf lors du processus de fécondation. DANS col de l'utérus sont les centrioles du centre cellulaire et la base queue de cheval forment des microtubules qui soutiennent directement le mouvement des spermatozoïdes. Il contient également des mitochondries, qui fournissent aux spermatozoïdes l’énergie ATP nécessaire au mouvement.
Oogenèse(du grec ONU- oeuf et genèse- origine, apparition) est le processus de formation de cellules germinales femelles matures - les œufs. Chez l'homme, elle se produit dans les ovaires et comprend trois périodes : la reproduction, la croissance et la maturation. Des périodes de reproduction et de croissance, similaires à celles de la spermatogenèse, se produisent au cours du développement intra-utérin. Dans ce cas, les cellules diploïdes sont formées à partir de cellules germinales primaires à la suite de la mitose. oogonie, qui se transforment ensuite en diploïdes primaires ovocytes, ou Ovules de 1er ordre. Méiose et cytokinèse ultérieure se produisant dans période de maturation, sont caractérisés par une division inégale du cytoplasme de la cellule mère, de sorte que l'on obtient d'abord un ovocyte secondaire, ou Ovocyte de 2ème ordre, Et premier corps polaire, puis de l'ovocyte secondaire - un ovule qui conserve la totalité de l'approvisionnement nutriments, et un deuxième globule polaire, tandis que le premier globule polaire est divisé en deux. Les corps polaires absorbent l’excès de matériel génétique.
Chez l'homme, les œufs sont produits à des intervalles de 28 à 29 jours. Le cycle associé à la maturation et à la libération des ovules est appelé menstruel.
Œuf- une grande cellule reproductrice femelle qui porte non seulement un ensemble haploïde de chromosomes, mais également un apport important de nutriments pour le développement ultérieur de l'embryon.

L'œuf des mammifères est recouvert de quatre membranes, ce qui réduit le risque de dommages causés par divers facteurs. Le diamètre de l'œuf chez l'homme atteint 150 à 200 microns, tandis que chez l'autruche, il peut atteindre plusieurs centimètres.
La division cellulaire est la base de la croissance, du développement et de la reproduction des organismes. Le rôle de la mitose et de la méiose
Si dans les organismes unicellulaires, la division cellulaire conduit à une augmentation du nombre d'individus, c'est-à-dire à la reproduction, alors dans les organismes multicellulaires, ce processus peut avoir sens différent. Ainsi, la division des cellules embryonnaires, à partir du zygote, constitue la base biologique des processus interconnectés de croissance et de développement. Des changements similaires sont observés chez l'homme au cours de l'adolescence, lorsque le nombre de cellules augmente non seulement, mais qu'un changement qualitatif se produit également dans le corps. La base de la reproduction des organismes multicellulaires est également la division cellulaire, par exemple, dans la reproduction asexuée, grâce à ce processus, toute une partie de l'organisme est restaurée, et dans la reproduction sexuée, au cours du processus de gamétogenèse, des cellules sexuelles se forment, qui donnent ensuite naissance à un nouvel organisme. Il convient de noter que les principales méthodes de division d'une cellule eucaryote - la mitose et la méiose - ont des significations différentes dans Les cycles de la vie organismes.
À la suite de la mitose, il existe une répartition uniforme du matériel héréditaire entre les cellules filles - copies exactes maternel. Sans mitose, l’existence et la croissance d’organismes multicellulaires se développant à partir d’une seule cellule, le zygote, seraient impossibles, puisque toutes les cellules de ces organismes doivent contenir la même information génétique.
Au cours du processus de division, les cellules filles deviennent de plus en plus diversifiées dans leur structure et leurs fonctions, ce qui est associé à l'activation de plus en plus de nouveaux groupes de gènes en raison de l'interaction intercellulaire. Ainsi, la mitose est nécessaire au développement de l’organisme.
Cette méthode de division cellulaire est nécessaire aux processus reproduction asexuée et la régénération (restauration) des tissus endommagés, ainsi que des organes.
La méiose, à son tour, assure la constance du caryotype lors de la reproduction sexuée, puisqu'elle divise par deux l'ensemble des chromosomes avant la reproduction sexuée, qui est ensuite restauré suite à la fécondation. De plus, la méiose conduit à l'émergence de nouvelles combinaisons de gènes parentaux dues au croisement et à la combinaison aléatoire de chromosomes dans les cellules filles. Grâce à cela, la progéniture s'avère génétiquement diversifiée, ce qui fournit du matériel pour sélection naturelle et constitue la base matérielle de l'évolution. Une modification du nombre, de la forme et de la taille des chromosomes, d'une part, peut conduire à l'apparition de diverses anomalies dans le développement de l'organisme et même à sa mort, et d'autre part, elle peut conduire à l'apparition d'individus. plus adapté à l'environnement.
Ainsi, la cellule est l’unité de croissance, de développement et de reproduction des organismes.
Chaque protéine est représentée par une ou plusieurs chaînes polypeptidiques. Une section d'ADN qui contient des informations sur une chaîne polypeptidique est appelée génome. La totalité des molécules d’ADN d’une cellule agit comme porteur d’informations génétiques. L'information génétique est transmise à la fois des cellules mères aux cellules filles et des parents aux enfants. Un gène est une unité génétique, ou informations héréditaires.
L'ADN est le porteur de l'information génétique dans une cellule– ne participe pas directement à la synthèse des protéines. Dans les cellules eucaryotes, les molécules d'ADN sont contenues dans les chromosomes du noyau et sont séparées par l'enveloppe nucléaire du cytoplasme, où se produit la synthèse des protéines. Un messager porteur d'informations est envoyé du noyau aux ribosomes, site d'assemblage des protéines, et est capable de traverser les pores de la membrane nucléaire. Ce messager est l’ARN messager (ARNm). Selon le principe de complémentarité, il est synthétisé sur l'ADN avec la participation d'une enzyme appelée ARN. polymérase.
1) Le processus de synthèse de l'ARN, dans lequel l'une des chaînes de la molécule d'ADN est utilisée comme matrice, est appelé transcription .
2) Transcription est un mécanisme par lequel la séquence nucléotidique de l'une des chaînes d'ADN est réécrite par la séquence complémentaire de la molécule d'ARNm.
L'ARN messager est une molécule simple brin et la transcription se produit à partir d'un brin d'une molécule d'ADN double brin. Il ne s'agit pas d'une copie de la molécule d'ADN entière, mais seulement d'une partie de celle-ci - un gène chez les eucaryotes ou un groupe de gènes adjacents qui portent des informations sur la structure des protéines nécessaires pour remplir une fonction chez les procaryotes. Ce groupe de gènes est appelé opéron. Au début de chaque opéron se trouve une sorte de plate-forme d'atterrissage pour l'ARN polymérase appelée promoteur.il s’agit d’une séquence spécifique de nucléotides d’ADN que l’enzyme « reconnaît » en raison de son affinité chimique. Ce n’est qu’en se fixant au promoteur que l’ARN polymérase est capable de commencer la synthèse de l’ARN. Ayant atteint la fin de l'opéron, l'enzyme rencontre un signal (sous la forme d'une certaine séquence nucléotidique) indiquant la fin de la lecture. L'ARNm fini quitte l'ADN et se dirige vers le site de synthèse des protéines.
Il y a quatre étapes dans le processus de transcription : 1) Liaison à l'ARN-polymérase avec un promoteur ; 2) initiation– début de synthèse. Elle consiste en la formation de la première liaison phosphodiester entre l'ATP ou le GTP et le deuxième nucléotide de la molécule d'ARN synthétisée ; 3) élongation– croissance de la chaîne d'ARN ; ceux. addition séquentielle de nucléotides les uns aux autres dans l’ordre dans lequel leurs nucléotides complémentaires apparaissent dans le brin d’ADN transcrit. Le taux d'élongation est de 50 nucléotides par seconde ; 4) Résiliation– achèvement de la synthèse d’ARN.
Après avoir traversé les pores de la membrane nucléaire, l'ARNm est envoyé aux ribosomes, où l'information génétique est déchiffrée - traduite du « langage » des nucléotides au « langage » des acides aminés. La synthèse de chaînes polypeptidiques à l'aide d'une matrice d'ARNm, qui se produit dans les ribosomes, est appelée diffuser(Traduction latine - traduction).
Les acides aminés, à partir desquels les protéines sont synthétisées, sont délivrés aux ribosomes à l'aide d'ARN spéciaux appelés ARN de transfert (ARNt). Il y a autant d’ARNt différents dans une cellule qu’il y a de codons codant pour des acides aminés. Au sommet de la « feuille » de chaque ARNt se trouve une séquence de trois nucléotides complémentaires des nucléotides du codon de l’ARNm. Ils l'appellent anticodon. Une enzyme spéciale, la codase, reconnaît l'ARNt et attache un acide aminé au « pétiole de la feuille » – uniquement celui codé par le triplet complémentaire de l'anticodon. La formation d’une liaison covalente entre l’ARNt et son « propre » acide aminé nécessite l’énergie d’une molécule d’ATP.
 Pour qu’un acide aminé soit inclus dans une chaîne polypeptidique, il doit se détacher de l’ARNt. Cela devient possible lorsque l’ARNt pénètre dans le ribosome et que l’anticodon reconnaît son codon dans l’ARNm. Le ribosome possède deux sites pour lier deux molécules d'ARNt. Dans l'une de ces zones, appelée accepteur, l'ARNt arrive avec un acide aminé et s'attache à son codon (I). Cet acide aminé s'attache-t-il (accepte-t-il) la chaîne protéique en croissance (II) ? Une liaison peptidique se forme entre eux. L'ARNt, qui est maintenant attaché avec le codon de l'ARNm dans donneur section du ribosome. Un nouvel ARNt arrive dans le site accepteur libéré, lié à un acide aminé, qui est crypté par le codon suivant (III). La chaîne polypeptidique détachée est ici à nouveau transférée du site donneur et étendue par un maillon supplémentaire. Les acides aminés de la chaîne en croissance sont connectés dans la séquence dans laquelle les codons les codant se trouvent dans l'ARNm.
Pour qu’un acide aminé soit inclus dans une chaîne polypeptidique, il doit se détacher de l’ARNt. Cela devient possible lorsque l’ARNt pénètre dans le ribosome et que l’anticodon reconnaît son codon dans l’ARNm. Le ribosome possède deux sites pour lier deux molécules d'ARNt. Dans l'une de ces zones, appelée accepteur, l'ARNt arrive avec un acide aminé et s'attache à son codon (I). Cet acide aminé s'attache-t-il (accepte-t-il) la chaîne protéique en croissance (II) ? Une liaison peptidique se forme entre eux. L'ARNt, qui est maintenant attaché avec le codon de l'ARNm dans donneur section du ribosome. Un nouvel ARNt arrive dans le site accepteur libéré, lié à un acide aminé, qui est crypté par le codon suivant (III). La chaîne polypeptidique détachée est ici à nouveau transférée du site donneur et étendue par un maillon supplémentaire. Les acides aminés de la chaîne en croissance sont connectés dans la séquence dans laquelle les codons les codant se trouvent dans l'ARNm.
Lorsqu'un des trois triplets apparaît sur le ribosome ( UAA, UAG, UGA), qui sont des « signes de ponctuation » entre les gènes, aucun ARNt ne peut prendre place dans le site accepteur. Le fait est qu'il n'existe pas d'anticodons complémentaires aux séquences nucléotidiques des « signes de ponctuation ». Le brin détaché n’a rien à quoi s’attacher au site accepteur et quitte le ribosome. La synthèse des protéines est terminée.
Chez les procaryotes, la synthèse des protéines commence par le codon AOÛT, situé en premier lieu dans la copie de chaque gène, occupe une position telle dans le ribosome que l'anticodon d'un ARNt spécial qui lui est connecté interagit avec formylmentionine. Cette forme modifiée de l'acide aminé méthionine pénètre immédiatement dans le site donneur et agit comme une majuscule dans la phrase - la synthèse de toute chaîne polypeptidique commence par elle dans la cellule bactérienne. Quand un triplet AOÛT n'est pas en premier lieu, mais à l'intérieur d'une copie du gène ; il code pour l'acide aminé méthionine. Une fois la synthèse de la chaîne polypeptidique terminée, la formylméthionine en est séparée et est absente de la protéine finie.
 Pour augmenter la production de protéines, l’ARNm traverse souvent non pas un mais plusieurs ribosomes en même temps. Cette structure, unie par une molécule d'ARNm, est appelée polysome. Chaque ribosome de ce tapis roulant en forme de billes synthétise les mêmes protéines.
Pour augmenter la production de protéines, l’ARNm traverse souvent non pas un mais plusieurs ribosomes en même temps. Cette structure, unie par une molécule d'ARNm, est appelée polysome. Chaque ribosome de ce tapis roulant en forme de billes synthétise les mêmes protéines.
Les acides aminés sont fournis en continu aux ribosomes à l’aide de l’ARNt. Après avoir fait don de l'acide aminé, l'ARNt quitte le ribosome et s'unit à l'aide de la codase. La grande cohérence de tous les « services de la plante » pour la production de protéines permet de synthétiser des chaînes polypeptidiques constituées de centaines d'acides aminés en quelques secondes.
Propriétés du code génétique. Grâce au processus de transcription dans la cellule, l'information est transférée de l'ADN aux protéines.
ADN → ARNm → protéine
L'information génétique contenue dans l'ADN et l'ARNm est contenue dans la séquence de nucléotides des molécules.
 Comment l’information est-elle transférée du « langage » des nucléotides au « langage » des acides aminés ? Cette traduction est réalisée à l'aide du code génétique. Code ou chiffre, est un système de symboles permettant de traduire une forme d'information en une autre. Code génétique est un système d'enregistrement d'informations sur la séquence d'acides aminés dans les protéines en utilisant la séquence de nucléotides dans l'ARNm.
Comment l’information est-elle transférée du « langage » des nucléotides au « langage » des acides aminés ? Cette traduction est réalisée à l'aide du code génétique. Code ou chiffre, est un système de symboles permettant de traduire une forme d'information en une autre. Code génétique est un système d'enregistrement d'informations sur la séquence d'acides aminés dans les protéines en utilisant la séquence de nucléotides dans l'ARNm.
Quelles propriétés possède le code génétique ?
1. Le code est un triplet. L'ARN contient quatre nucléotides : A, G, C, U. Si nous essayions de désigner un acide aminé avec un nucléotide, alors 16 des 20 acides aminés resteraient non codés. Un code à deux lettres chiffrerait 16 acides aminés. La nature a créé un code à trois lettres, ou triplet. Cela signifie que Chacun des 20 acides aminés est codé par une séquence de trois nucléotides appelée triplet ou codon.
2. Le code est dégénéré. Cela signifie que Chaque acide aminé est codé par plusieurs codons. Exceptions : la météonine et le tryptophane, chacun étant codé par un triplet.
3. Le code est clair. Chaque codon code pour un seul acide aminé.
4. Il y a des « signes de ponctuation » entre les gènes. Dans le texte imprimé, il y a un point à la fin de chaque phrase. Plusieurs phrases liées composent un paragraphe. Dans le langage de l'information génétique, un tel paragraphe est un opéron et son ARNm complémentaire. Chaque gène d'un opéron procaryote ou d'un gène eucaryote distinct code pour une chaîne polypeptidique - une phrase. Étant donné que dans certains cas, plusieurs chaînes polypeptidiques différentes sont créées séquentiellement à partir de la matrice d'ARNm, elles doivent être séparées les unes des autres. À cette fin, il existe trois triplets spéciaux dans l'année génétique - UAA, UAG, UGA, dont chacun indique l'arrêt de la synthèse d'une chaîne polypeptidique. Ainsi, ces triplets fonctionnent comme des signes de ponctuation. On les trouve à la fin de chaque gène.
5. Il n’y a pas de « signes de ponctuation » dans un gène.
6. Le code est universel. Le code génétique est le même pour toutes les créatures vivant sur Terre. Chez les bactéries et les champignons, le blé et le coton, les poissons et les vers, les grenouilles et les humains, les mêmes triplets codent pour les mêmes acides aminés.
 Principes de réplication de l'ADN.
La continuité du matériel génétique dans les générations de cellules et d'organismes est assurée par le processus réplication - doublement des molécules d'ADN. Ce processus complexe est réalisé par un complexe de plusieurs enzymes et protéines sans activité catalytique, nécessaires pour donner aux chaînes polynucléotidiques la conformation souhaitée. À la suite de la réplication, deux doubles hélices d'ADN identiques se forment. Ces molécules dites filles ne sont pas différentes les unes des autres ni de la molécule d’ADN mère d’origine. La réplication se produit dans la cellule avant la division, de sorte que chaque cellule fille reçoit exactement les mêmes molécules d'ADN que la cellule mère. Le processus de réplication repose sur un certain nombre de principes :
Principes de réplication de l'ADN.
La continuité du matériel génétique dans les générations de cellules et d'organismes est assurée par le processus réplication - doublement des molécules d'ADN. Ce processus complexe est réalisé par un complexe de plusieurs enzymes et protéines sans activité catalytique, nécessaires pour donner aux chaînes polynucléotidiques la conformation souhaitée. À la suite de la réplication, deux doubles hélices d'ADN identiques se forment. Ces molécules dites filles ne sont pas différentes les unes des autres ni de la molécule d’ADN mère d’origine. La réplication se produit dans la cellule avant la division, de sorte que chaque cellule fille reçoit exactement les mêmes molécules d'ADN que la cellule mère. Le processus de réplication repose sur un certain nombre de principes :
1. Complémentarité. Chacune des deux chaînes de la molécule d'ADN parent sert de modèle pour la synthèse de son complément, c'est-à-dire complémentaire, chaîne fille.
2. Semi-conservateur. À la suite de la réplication, deux hélices filles doubles se forment, chacune conservant (préservant) l'un des brins d'ADN parent inchangé. Les secondes chaînes de molécules filles sont à nouveau synthétisées à partir de nucléotides selon la complémentarité avec les brins d'ADN maternel. Les ADN filles ne sont pas différents les uns des autres ni de la double hélice parentale.
3.
 Antiparallélisme. Chaque brin d'ADN a une orientation spécifique. Une extrémité porte un groupe hydroxyle (-OH) attaché au carbone 3' du sucre désoxyribose, et l'autre extrémité de la chaîne a un résidu d'acide phosphorique en position 5' du sucre. Les deux brins complémentaires d’une molécule d’ADN sont orientés dans des directions opposées : antiparallèles. Les enzymes qui synthétisent de nouveaux brins d'ADN et sont appelées ADN polymérases peuvent se déplacer le long des brins matrices dans une seule direction : de leurs extrémités 3' à leurs extrémités 5'. Dans ce cas, la synthèse des brins complémentaires s’effectue toujours dans le sens 5’ → 3’, c’est-à-dire unipolaire. Par conséquent, au cours du processus de réplication, la synthèse simultanée de nouvelles chaînes se produit de manière antiparallèle.
Antiparallélisme. Chaque brin d'ADN a une orientation spécifique. Une extrémité porte un groupe hydroxyle (-OH) attaché au carbone 3' du sucre désoxyribose, et l'autre extrémité de la chaîne a un résidu d'acide phosphorique en position 5' du sucre. Les deux brins complémentaires d’une molécule d’ADN sont orientés dans des directions opposées : antiparallèles. Les enzymes qui synthétisent de nouveaux brins d'ADN et sont appelées ADN polymérases peuvent se déplacer le long des brins matrices dans une seule direction : de leurs extrémités 3' à leurs extrémités 5'. Dans ce cas, la synthèse des brins complémentaires s’effectue toujours dans le sens 5’ → 3’, c’est-à-dire unipolaire. Par conséquent, au cours du processus de réplication, la synthèse simultanée de nouvelles chaînes se produit de manière antiparallèle.
4. Discontinuité. Pour que de nouveaux brins d’ADN soient construits selon le principe de complémentarité, la double hélice doit être déroulée et il ne doit y avoir aucune liaison hydrogène entre les brins mères.

 Ce n'est que dans ce cas que les ADN polymérases sont capables de se déplacer le long des brins mères et de les utiliser comme modèles pour la synthèse sans erreur des brins filles. Mais le déroulement complet d'hélices constituées de plusieurs millions de paires de nucléotides est associé à un nombre de rotations si important et à des coûts énergétiques tels qu'ils sont impossibles dans des conditions cellulaires. Par conséquent, la réplication chez les eucaryotes commence simultanément à certains endroits de la molécule d'ADN. La zone située entre les deux points où commence la synthèse des chaînes filles est appelée réplicon. Il est unité de réplication.
Ce n'est que dans ce cas que les ADN polymérases sont capables de se déplacer le long des brins mères et de les utiliser comme modèles pour la synthèse sans erreur des brins filles. Mais le déroulement complet d'hélices constituées de plusieurs millions de paires de nucléotides est associé à un nombre de rotations si important et à des coûts énergétiques tels qu'ils sont impossibles dans des conditions cellulaires. Par conséquent, la réplication chez les eucaryotes commence simultanément à certains endroits de la molécule d'ADN. La zone située entre les deux points où commence la synthèse des chaînes filles est appelée réplicon. Il est unité de réplication.
Chaque molécule d'ADN d'une cellule eucaryote contient de nombreux réplicons. Dans chaque réplicon, vous pouvez voir une fourche de réplication - cette partie de la molécule d'ADN qui s'est déjà détachée sous l'influence d'enzymes spéciales. Chaque brin de la fourche sert de modèle pour la synthèse d'un brin fille complémentaire. Lors de la réplication, la fourche se déplace le long de la molécule mère et de nouvelles sections d'ADN se déroulent. Étant donné que les ADN polymérases ne peuvent se déplacer que dans une seule direction le long des brins matrices et que les brins sont orientés de manière antiparallèle, deux complexes enzymatiques différents sont synthétisés simultanément dans chaque fourche. De plus, dans chaque fourche, une chaîne fille (principale) croît continuellement, tandis que l'autre chaîne (en retard) est synthétisée en fragments séparés longs de plusieurs nucléotides. Ces enzymes, du nom du scientifique japonais qui les a découvertes fragments d'Okazaki, sont réticulés avec l'ADN ligase pour former une chaîne continue. Le mécanisme de formation des brins d'ADN filles par fragments est dit discontinu.
5. La nécessité d'amorçage de l'ADN polymérase est incapable d'initier la synthèse du brin leader, ni la synthèse des fragments d'Okazaki du brin retardateur. Il ne peut étendre un brin polynucléotidique existant qu’en ajoutant séquentiellement des désoxyribonucléotides à son extrémité 3’-OH. D'où vient la région terminale 5' initiale de la chaîne d'ADN en croissance ? Il est synthétisé sur une matrice d'ADN par une ARN polymérase spéciale appelée prime(Anglais Primer - graine). La taille de l'amorce ribonucléotidique est petite (moins de 20 nucléotides) par rapport à la taille de la chaîne d'ADN formée par l'ADN poimérase. L'ayant complétée Fonction L'amorce ARN est éliminée par une enzyme spéciale et l'espace formé au cours de ce processus est comblé par l'ADN polymérase, qui utilise l'extrémité 3'-OH du fragment Okazaki adjacent comme amorce.
Le problème de la sous-réplication des extrémités des molécules d'ADN linéaires. L'élimination des amorces d'ARN extrêmes, complémentaires des extrémités 3' des deux brins de la molécule d'ADN parent linéaire, donne lieu à des brins filles plus courts que 10 à 20 nucléotides. C’est le problème de la sous-réplication des extrémités des molécules linéaires.
Le problème de la sous-réplication des extrémités 3' des molécules d'ADN linéaires est résolu par les cellules eucaryotes à l'aide d'une enzyme spéciale - télomérase.
La télomérase est une ADN polymérase qui complète les molécules d'ADN 3'-terminales des chromosomes avec de courtes séquences répétitives. Ils, situés les uns derrière les autres, forment une structure terminale régulière pouvant atteindre 10 000 nucléotides de long. En plus de la partie protéique, la télomérase contient de l'ARN, qui sert de modèle pour étendre les répétitions d'ADN.
Schéma d'allongement des extrémités des molécules d'ADN. Tout d’abord, une liaison complémentaire de l’extrémité saillante de l’ADN à la région matrice de l’ARN de la télomérase se produit, puis la télomérase étend l’ADN en utilisant son extrémité 3’-OH comme amorce et l’ARN inclus dans l’enzyme comme matrice. Cette étape est appelée allongement. Après cela, la translocation se produit, c'est-à-dire mouvement de l'ADN prolongé d'une répétition par rapport à l'enzyme. Ceci est suivi d'un allongement et d'une autre translocation.
En conséquence, des structures terminales chromosomiques spécialisées se forment. Ils sont constitués de courtes séquences d’ADN et de protéines spécifiques répétées à plusieurs reprises.
Bref résumé.
Déterminer la possibilité de développer un trait particulier
d'une cellule ou d'un organisme est un GÈNE.
Lorsque les gènes sont transmis sur plusieurs générations,
héritage des traits parentaux par la progéniture.
La propriété principale du gène en tant qu'unité fonctionnelle
matière d'hérédité et de variabilité, est son
organisation chimique.
Lors de la formation de fonctionnalités, il est nécessaire
synthèse de nombreuses substances, d'une part
séquence de protéines avec des
propriétés.
Propriété
protéine
la molécule est déterminée par l'acide aminé
séquence
son
peptide
Chaînes,
lequel
est donné
séquence de nucléotides d’ADN.
Acides nucléiques - ADN et ARN
Acides nucléiques ADN et ARNIl existe deux types d'acides nucléiques
Désoxyribonucléiqueacide (ADN), inclus
qui contient le désoxyribose glucidique
Ribonucléique
acide (ARN), inclus
qui contient le glucide ribose. L'ADN est la plus grosse molécule d'une cellule.
C'est beaucoup plus gros que les protéines et l'ARN
Chaque chromosome = une molécule d'ADN
23 chromosomes humains = 23 molécules d'ADN
Les plus longs d'entre eux mesurent ≈ 8 cm
L'ADN est une molécule de texte. DANS
ses séquences nucléotidiques
tout le programme héréditaire est enregistré
corps
Localisation de l'ADN dans une cellule
CœurMitochondries
Plastides
Cœur
Chloroplaste
Mitochondries 1 molécule d'ADN
gène
un autre gène
chromosome
chromosomes dans
cœur
ADN
cellule
Fonctions de l'ADN
Stockagegénétique
information
Diffuser
génétique
Information provenant de
parents
postérité
Mise en œuvre
génétique
informations dans
processus
vitalement actif
cellules sti et
corps
Consolidation
Histoire de la découverte
1. 1869 Friedrich Miescherdécouvert NK et leur a donné
nom (noyau « noyau »).
2. 1905 Edwin Chargaff
nucléotide étudié
composition de NK.
3. 1950 Rosalinde
Franklin a établi
ADN double brin.
Edwin
Chargaff
Rosalinde
Franklin Diffraction des rayons X
Portrait ADN –
célèbre photo 51
Rosalinde Franklin
1920 - 1958http://www.bbc.co.uk/bbcfour/documentaries/features/rosalind- 1953
Ouvrir
structure
ADN
date
naissance
moléculaire
la biologie
James
Watson
Francis
Crier James Dewey
Watson
Francis Harry Compton
Crampe
Les molécules d'ADN peuvent être vues au microscope électronique
ADN de plasmides bactériens ADN du réovirusbalayage électrique microscope ADN isolé
d'un chromosome
personne
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/L/Laemmli
L'ADN est une molécule polymère constituée de 2 chaînes polynucléotidiques complémentaires reliées par des liaisons hydrogène, elles ont de grandes
Structure de la molécule d'ADNL'ADN est un polymère
molécule composée de 2
gratuit
chaînes polynucléotidiques
connecté
hydrogène
Connexions,
sont de grande taille et
énorme molécule
masse.
Structure de la molécule d'ADN
Chaînes nucléotidiquesformulaire
droitier
spirales volumétriques de 10 paires
bases à chaque tour
Les chaînes s'enroulent
les uns les autres et autour
axe et forme communs
double hélice
Les circuits sont antiparallèles ou
multidirectionnel.
Sous-séquence
composés nucléotidiques
une chaîne
à l'opposé de celui dans
un autre
Structure schématique de l'ADN
Nucléotides :1. Situés à l’écart les uns des autres
ami à distance
0,34 nm
2. Masse d'un
le nucléotide est 345.
3. Largeur d'hélice 2 nm
4. Ces valeurs
permanent Principes de structure de l'ADN
5"
3"
UN
g
g
T
C
UN
UN
C
5"
Irrégularité
T
C
C
UN
Double brin
Complémentarité
Antiparallélisme
g
T
T
g
3"
Structure de l'ADN
L'ADN est un polymère.Les monomères sont des nucléotides.
Nucléotide - composé chimique
résidus de trois substances :
Structure nucléotidique
Azoté
terrains:
- Adénine ;
- Guanine ;
- Cytazine
- Timin
Glucides :
- Désoxyribose
Résidu de phosphore
acides (AF) Nucléotide
5’
1’
phosphate
Azoté
base -
un des 4
3’
Sucre (ribose/désoxyribose)
Schémas de la structure des bases azotées.
L'ADN contientsuivant
azoté
terrains:
Purine
1. Adénine,
2. Guanine
Pyrimidine
3. Timine
4. Cytazine
Connexions entre les nucléotides dans un brin d'ADN
Sont en courspar l'éducation
phosphoester
les liens entre
désoxyribose un
nucléotide et résidu
acide phosphorique
un autre nucléotide
Liens entre brins dans une molécule d'ADN
Mis en œuvreAvec de l'aide
les liaisons hydrogène,
émergent
entre
azoté
les raisons
boîte de réception
V
composé
différents circuits
Règles des Chargaffs de 1950
Erwin ChargaffLes règles de Chargaff ont été expliquées par Watson et Crick
L'ADN est constitué de 2 brins connectésselon le principe
complémentarité
Complémentarité
ComplémentaritéCe
principe de mutuelle
correspondances ou capacités de nucléotides appariés
les nucléotides se combinent par paires
Règles de la Chargaffe
[A] + [G] = [T] + [C] = 50 % Principecomplémentarité :
UN
-- -- --
T
g
-- -- ----
C
Plus durable
Faible
hydrogène
Connexions!
Principe de complémentarité
Propriété de "réplication"
La réplication de l'ADN estprocessus de copie
acide désoxyribonucléique
acide, qui
se produit dans le processus
la division cellulaire.
En même temps, la génétique
matériel crypté
dans l'ADN, double et
réparti entre filiales
cellules.
Réplication de l'ADN
Danstemps
réplication
Partie
molécules
L'ADN « maternel » se déroule en deux brins avec
en utilisant une enzyme spéciale, et ce
obtenu en brisant les liaisons hydrogène entre
bases azotées complémentaires :
adénine-thymine et guanine-cytosine.
Suite à chaque nucléotide des fils divergents
L'ADN enzyme ADN polymérase s'ajuste
son nucléotide complémentaire.
Propriété de "réplication"
Code génétique
Les informations héréditaires sont enregistrées dansMolécules NA sous forme de séquence
nucléotides.
Certain
parcelles
Molécules d'ADN et d'ARN (dans les virus et les phages)
contenir des informations sur la structure principale
une protéine et sont appelés gènes.
1 gène = 1 molécule de protéine
C'est pourquoi
héréditaire
information,
lequel
contenir
ADN
appelé
génétique.
Propriétés du code génétique :
PolyvalenceDiscrétion
(code
triplés
lire à partir de la molécule d'ARN entière)
Spécificité (le codon code uniquement pour AK)
Redondance du code (un peu)
Propriété de « réparation »
La réparation est la capacité d'une molécule d'ADNcorriger ceux qui se produisent dans ses circuits
changements.
En restaurant la structure originale de l'ADN
Au moins 20 protéines sont impliquées :
1. Reconnaître les sections d'ADN modifiées ;
2. Retirez-les du circuit ;
3. Restaurez le bon
séquence nucléotidique;
4. Cousez le fragment restauré avec
le reste de la molécule d'ADN
Bibliographie
Zakharov V.B. et etc. " Biologie générale”Ruvinsky A.O. Moscou « Lumières »
1993 « Biologie générale »
«La biologie en tableaux et diagrammes»,
«Outarde» 2005
Internet : Google
Acide désoxyribonucléique(ADN) est un support matériel d’information génétique. Il s'agit d'un composé naturel de haut poids moléculaire contenu dans les noyaux des cellules des organismes vivants. Les molécules d'ADN et les protéines histones forment une substance chromosomes. Les histones font partie des noyaux cellulaires et participent au maintien et à la modification de la structure des chromosomes à différentes étapes du cycle cellulaire et à la régulation de l'activité des gènes. Des sections individuelles de molécules d'ADN correspondent à des gènes spécifiques. Une molécule d'ADN est constituée de deux chaînes polynucléotidiques enroulées l'une autour de l'autre en spirale (Fig. 7.1). Les chaînes sont constituées d'un grand nombre de monomères de quatre types - les nucléotides, dont la spécificité est déterminée par l'une des quatre bases azotées : adénine(UN), thymine(T), cytosine(C) et guanine(G). La combinaison de trois nucléotides adjacents dans une chaîne d'ADN forme code génétique. La violation de la séquence nucléotidique dans la chaîne d'ADN entraîne des modifications héréditaires dans le corps - mutations. L'ADN est reproduit avec précision lors de la division cellulaire, ce qui assure la transmission des caractéristiques héréditaires et des formes spécifiques de métabolisme sur une série de générations de cellules et d'organismes.
Riz. 7.1. Structure d'une molécule d'ADN.
Le modèle structurel de l'ADN en forme de double hélice a été proposé en 1953 par le biochimiste américain J. Watson (né en 1928) et le biophysicien et généticien anglais F. Crick (né en 1916). Le modèle Watson-Crick a permis d'expliquer de nombreuses propriétés et fonctions biologiques de la molécule d'ADN. Pour avoir déchiffré le code génétique, J. Watson, F. Crick et le biophysicien anglais M. Wilkins (né en 1916), qui fut le premier à obtenir une photographie aux rayons X de haute qualité d'une molécule d'ADN, ont été récompensés. prix Nobel 1962
L'ADN est une formation naturelle étonnante avec une symétrie spirale. Les longs brins entrelacés de la structure de la chaîne d’ADN sont constitués de molécules de sucre et de phosphate. Les bases azotées sont attachées aux molécules de sucre, formant des liaisons croisées entre les deux brins hélicoïdaux. Une molécule d’ADN allongée ressemble à un escalier en colimaçon déformé. Il s'agit véritablement d'une macromolécule : son masse moléculaire peut atteindre 10 9. Malgré sa structure complexe, la molécule d'ADN ne contient que quatre bases azotées : A, T, C, G. Des liaisons hydrogène se forment entre l'adénine et la thymine. Ils sont si structurellement cohérents les uns avec les autres que l'adénine reconnaît et se lie à la thymine, et vice versa. La cytosine et la guanine forment une autre paire d’un type similaire. Dans ces paires de nucléotides, A est donc toujours associé à T, et C à G (Fig. 7.2). Cette connexion correspond le principe de complémentarité. Le nombre de paires de bases : adénine-thymine et cytosine-guanine par exemple, chez l'homme est énorme : certains chercheurs estiment qu'il y en a 3 milliards, tandis que d'autres estiment qu'il y en a plus de 3,5 milliards.

La capacité des bases azotées à reconnaître leur partenaire conduit au repliement de chaînes sucre-phosphate sous la forme d'une double hélice dont la structure a été déterminée expérimentalement grâce à des observations aux rayons X. Les interactions entre les bases azotées sont très spécifiques, donc une hélice ne peut se former que si les séquences de bases des deux brins sont complètement identiques.
Un groupe sucre phosphate avec l'une des bases azotées A, T, C ou G, formant nucléotide(Fig. 7.3) peut être représenté comme une sorte d’élément de base. La molécule d'ADN est constituée de tels blocs. La séquence de nucléotides code des informations dans une molécule d'ADN. Il contient des informations nécessaires, par exemple, à la production de protéines nécessaires à un organisme vivant.

Une molécule d'ADN peut être copiée dans un processus catalysé par des enzymes réplication, ce qui consiste à le doubler. Lors de la réplication, les liaisons hydrogène sont rompues pour former des chaînes simples qui servent de modèle pour la synthèse enzymatique des mêmes séquences de blocs de construction. Le processus de réplication implique donc la rupture des anciennes liaisons hydrogène et la formation de nouvelles. Au début de la réplication, deux brins opposés commencent à se dérouler et à se séparer l'un de l'autre (Fig. 7.4). Au moment du déroulement, l'enzyme attache de nouvelles chaînes à deux anciennes selon le principe de complémentarité : T dans la nouvelle chaîne est situé en face de A dans l'ancienne, etc., de ce fait, deux doubles hélices identiques se forment. En raison de la relative faiblesse de ces liaisons, la réplication se produit sans rompre les liaisons covalentes plus fortes dans les chaînes sucre-phosphate. Le codage de l'information génétique et la réplication de la molécule d'ADN sont des processus essentiels interconnectés nécessaires au développement d'un organisme vivant.

L'information génétique est codée par la séquence de nucléotides d'ADN. Des travaux fondamentaux sur le déchiffrement du code génétique ont été réalisés par les biochimistes américains M. Nirenberg (né en 1927), X. Korana (né en 1922) et R. Holley (né en 1922) ; Lauréats du prix Nobel 1968 Trois nucléotides consécutifs constituent une unité de code génétique appelée codon. Chaque codon code pour un acide aminé particulier. nombre total dont il y en a 20. Une molécule d'ADN peut être représentée comme une séquence de lettres-nucléotides qui forment un texte à partir d'un grand nombre d'entre elles, par exemple, ASAT-TGGAG... Un tel texte contient des informations qui déterminent les spécificités de chacun organisme : une personne, un dauphin, etc. Le code génétique de tous les êtres vivants, qu'il s'agisse d'une plante, d'un animal ou d'une bactérie, est le même. Par exemple, le codon GGU code pour l’acide aminé glycine dans tous les organismes. Cette caractéristique du code génétique, ainsi que la similitude de la composition en acides aminés de toutes les protéines, indiquent l'unité biochimique de la vie, qui reflète apparemment l'origine de tous les êtres vivants d'un seul ancêtre.
1) ARNm 2) ARNt 3) ADN 4) chromosome
A2. Lors de leur reproduction, les cellules filles de la peau humaine reçoivent de la cellule mère :
informations génétiques complètes
la moitié des informations
un quart des informations
il n'y a pas de bonne réponse
A3. La réplication de l'ADN s'accompagne de la rupture des liaisons chimiques :
peptide, entre acides aminés
covalent, entre glucides et phosphates
hydrogène, entre bases azotées
ionique, dans la structure d'une molécule
A4. Lorsqu’une molécule d’ADN est répliquée, elle produit :
un fil qui s'est brisé en fragments séparés de molécules filles
molécule composée de deux nouveaux brins d'ADN
molécule dont la moitié est constituée d’un brin d’ARNm
molécule fille constituée d'un ancien et d'un nouveau brin d'ADN
A5. La transcription est le processus de :
1) Réplication de l'ADN
2) synthèse de l'ARNm
3) synthèse des protéines
4) joindre l'ARNt à l'acide aminé
A6. Si un acide aminé est codé par le codon UGG, alors dans l'ADN il correspond à un triplet :
TCC 2) AGG 3) UCC 4) ACC
A7. Un triplet d’ADN contient des informations sur :
Séquences d'acides aminés dans une molécule de protéine
La localisation d'un acide aminé spécifique dans une chaîne protéique
Signe d'un organisme spécifique
Un acide aminé inclus dans une chaîne protéique
A8. Le nombre d’ARNt impliqués dans la traduction est égal au nombre de :
Codons d'ARNm qui codent pour les acides aminés
Molécules d'ARNm
Gènes inclus dans une molécule d'ADN
Protéines synthétisées sur les ribosomes
A9. La période de vie d'une cellule, de division en division, s'appelle :
Interphase 3) méiose
Mitose 4) cycle cellulaire
A10. Combien de chromatides sont contenues dans les 8 chromosomes visibles en métaphase de mitose :
1) 6 2) 8 3) 12 4) 16
A11. Le nombre de chromosomes dans les cellules somatiques humaines après la mitose est :
1) 23 2) 46 3) 92 4) 44




