DNA on lühidalt päriliku teabe kandja. A1
Geneetiline teave rakus. Geenid, geneetiline kood ja selle omadused. Biosünteetiliste reaktsioonide maatriks olemus. Valkude ja nukleiinhapete biosüntees
Geneetiline teave rakus
Omasuguste paljunemine on elavate inimeste üks põhiomadusi. Selle nähtuse tõttu on sarnasus mitte ainult organismide, vaid ka üksikute rakkude, aga ka nende organellide (mitokondrid ja plastiidid) vahel. Selle sarnasuse materiaalseks aluseks on DNA nukleotiidjärjestuses krüpteeritud geneetilise teabe edastamine, mis toimub tänu DNA replikatsiooni protsessidele (isedubleerumine). Kõik rakkude ja organismide omadused ja omadused realiseeruvad tänu valkudele, mille struktuuri määrab eelkõige DNA nukleotiidide järjestus. Seetõttu on biosüntees ainevahetusprotsessides ülimalt tähtis. nukleiinhapped ja orav. Päriliku informatsiooni struktuuriüksus on geen.
Geenid, geneetiline kood ja selle omadused
Pärilik teave rakus ei ole monoliitne, see on jagatud eraldi "sõnadeks" - geenideks.
Gene on geneetilise informatsiooni põhiühik.
Mitmes riigis samaaegselt läbiviidud ja selle sajandi alguses valminud töö "Inimese genoomi" programmi kallal andis meile arusaama, et inimesel on vaid umbes 25-30 tuhat geeni, kuid info enamikust meie DNA-d ei loeta kunagi, sest see sisaldab suur summa mõttetud lõigud, kordused ja inimese jaoks tähenduse kaotanud tunnuseid kodeerivad geenid (saba, kehakarvad jne). Lisaks on dešifreeritud hulk geene, mis vastutavad pärilike haiguste tekke eest, samuti ravimite sihtgeenid. Kuid praktiline kasutamine selle programmi rakendamisel saadud tulemused lükatakse edasi seni, kuni enamate inimeste genoomid on lahti mõtestatud ja selgub, mille poolest need erinevad.
Nimetatakse geene, mis kodeerivad valgu primaarset struktuuri, ribosomaalset või ülekande-RNA-d struktuurne ja geenid, mis aktiveerivad või pärsivad struktuurigeenide teabe lugemist - regulatiivsed. Kuid isegi struktuursed geenid sisaldavad reguleerivaid piirkondi.
Organismide pärilik teave krüpteeritakse DNA-s teatud nukleotiidide kombinatsioonide ja nende järjestuse kujul - geneetiline kood. Selle omadused on: kolmik, spetsiifilisus, universaalsus, liiasus ja mittekattuvus. Lisaks puuduvad geneetilises koodis kirjavahemärgid.
Iga aminohapet kodeerib DNA kolm nukleotiidi - kolmik näiteks metioniini kodeerib TAC-triplet, see tähendab tripleti kood. Teisest küljest kodeerib iga kolmik ainult ühte aminohapet, mis on selle spetsiifilisus või ühemõttelisus. Geneetiline kood universaalne kõigile elusorganismidele, st pärilikku teavet inimese valkude kohta saavad lugeda bakterid ja vastupidi. See annab tunnistust päritolu ühtsusest orgaaniline maailm. Kolme nukleotiidi 64 kombinatsioonile vastavad aga vaid 20 aminohapet, mille tulemusena saavad ühte aminohapet kodeerida 2-6 tripletti ehk geneetiline kood on üleliigne ehk degenereerunud. Kolmel kolmikul pole vastavaid aminohappeid, neid nimetatakse stoppkoodonid, kuna need tähistavad polüpeptiidahela sünteesi lõppu.
Aluste järjestus DNA kolmikutes ja aminohapped, mida need kodeerivad
*Stoppkoodon, mis näitab polüpeptiidahela sünteesi lõppu.
Aminohapete nimetuste lühendid:
Ala - alaniin
Arg - arginiin
Asn - asparagiin
Asp - asparagiinhape
Val - valiin
Tema - histidiin
Gly - glütsiin
Gln – glutamiin
Glu - glutamiinhape
Ile - isoleutsiin
Leu - leutsiin
Liz - lüsiin
Meth - metioniin
Pro - proliin
Ser - seriin
Tyr - türosiin
Tre - treoniin
Kolm - trüptofaan
Fen - fenüülalaniin
cis - tsüsteiin
Kui hakkate geneetilist teavet lugema mitte tripleti esimesest nukleotiidist, vaid teisest, siis mitte ainult lugemisraam ei nihku - sel viisil sünteesitud valk on täiesti erinev mitte ainult nukleotiidjärjestuses, vaid ka struktuuris. ja omadused. Kolmikute vahel pole kirjavahemärke, seega pole lugemisraami nihutamisel takistusi, mis avab võimaluse mutatsioonide tekkeks ja säilimiseks.
Biosünteetiliste reaktsioonide maatriks olemus
Bakterirakud on võimelised dubleerima iga 20–30 minuti järel, eukarüootsed rakud aga iga päev ja veelgi sagedamini, mis nõuab DNA replikatsiooni suurt kiirust ja täpsust. Lisaks sisaldab iga rakk sadu ja tuhandeid koopiaid paljudest valkudest, eriti ensüümidest, seetõttu on nende paljundamiseks vastuvõetamatu nende tootmise "tükk" meetod. Progressiivsem viis on tembeldamine, mis võimaldab saada tootest arvukalt täpseid koopiaid ja ühtlasi vähendada selle maksumust. Tembeldamiseks on vaja maatriksit, millega tehakse jäljend.
Rakkudes on maatriksisünteesi põhimõte see, et uued valkude ja nukleiinhapete molekulid sünteesitakse vastavalt programmile, mis on sätestatud samade nukleiinhapete (DNA või RNA) olemasolevate molekulide struktuuris.
Valkude ja nukleiinhapete biosüntees
DNA replikatsioon. DNA on kaheahelaline biopolümeer, mille monomeerideks on nukleotiidid. Kui DNA biosüntees toimuks fotokopeerimise põhimõttel, tekiks paratamatult arvukalt moonutusi ja tõrkeid pärilikus informatsioonis, mis lõppkokkuvõttes tooks kaasa uute organismide hukkumise. Seetõttu on DNA dubleerimise protsess erinev, poolkonservatiivsel viisil: DNA molekul rullub lahti ja igal ahelal sünteesitakse komplementaarsuse põhimõttel uus ahel. DNA molekuli isepaljunemise protsessi, mis tagab päriliku informatsiooni täpse kopeerimise ja edasikandmise põlvest põlve nimetatakse nn. replikatsioon(alates lat. replikatsioon- kordamine). Replikatsiooni tulemusena moodustub DNA algmolekulist kaks absoluutselt täpset koopiat, millest igaüks kannab ühte algmolekuli koopiat.

Replikatsiooniprotsess on tegelikult äärmiselt keeruline, kuna selles osalevad mitmed valgud. Mõned neist kerivad lahti DNA kaksikheeliksi, teised lõhuvad komplementaarsete ahelate nukleotiidide vahelisi vesiniksidemeid, teised (näiteks DNA polümeraasi ensüüm) valivad komplementaarsuse põhimõttel uued nukleotiidid jne. Kaks DNA molekuli moodustuvad replikatsiooni tulemusena jagunevad jagunemise käigus kaheks vastloodud tütarrakud.
Vead replikatsiooniprotsessis on äärmiselt haruldased, kuid kui need ilmnevad, kõrvaldatakse need väga kiiresti nii DNA polümeraaside kui ka spetsiaalsete parandusensüümide poolt, kuna iga nukleotiidjärjestuse viga võib põhjustada valgu struktuuri ja funktsioonide pöördumatuid muutusi. ja lõppkokkuvõttes kahjustab uue raku või isegi indiviidi elujõulisust.
valkude biosüntees. Nagu 19. sajandi silmapaistev filosoof F. Engels piltlikult ütles: "Elu on valgukehade eksisteerimise vorm." Valgumolekulide struktuuri ja omadused määrab nende esmane struktuur, st DNA-s kodeeritud aminohapete järjestus. Selle teabe reprodutseerimise täpsusest ei sõltu mitte ainult polüpeptiidi enda olemasolu, vaid ka raku kui terviku toimimine, seetõttu on valkude sünteesi protsessil suur tähtsus. See näib olevat kõige keerulisem sünteesiprotsess rakus, kuna siin osaleb kuni kolmsada erinevat ensüümi ja muud makromolekuli. Lisaks töötab see koos suur kiirus mis nõuab veelgi suuremat täpsust.
Valkude biosünteesis on kaks peamist etappi: transkriptsioon ja translatsioon.
Transkriptsioon(alates lat. transkriptsioon- ümberkirjutamine) on mRNA molekulide biosüntees DNA matriitsil.

Kuna DNA molekul sisaldab kahte antiparalleelset ahelat, siis mõlemast ahelast info lugemine tooks kaasa täiesti erinevate mRNA-de moodustumise, mistõttu on nende biosüntees võimalik ainult ühes ahelas, mida nimetatakse kodeerivaks ehk kodogeenseks, erinevalt teisest. mittekodeerivad või mittekodogeensed. Ümberkirjutamise protsessi tagab spetsiaalne ensüüm RNA polümeraas, mis selekteerib RNA nukleotiide vastavalt komplementaarsuse põhimõttele. See protsess võib toimuda nii tuumas kui ka organellides, millel on oma DNA – mitokondrites ja plastiidides.
Transkriptsiooni käigus sünteesitud mRNA molekulid läbivad keerulise translatsiooni ettevalmistamise protsessi (mitokondriaalsed ja plastiidsed mRNA-d võivad jääda organellidesse, kus toimub valkude biosünteesi teine etapp). MRNA küpsemise protsessis kinnituvad sellele kolm esimest nukleotiidi (AUG) ja adenüülnukleotiidide saba, mille pikkus määrab, mitu valgu koopiat saab antud molekulil sünteesida. Alles siis lahkuvad küpsed mRNA-d tuumast läbi tuumapooride.
Paralleelselt toimub tsütoplasmas aminohapete aktiveerimise protsess, mille käigus seondub aminohape vastava vaba tRNA-ga. Seda protsessi katalüüsib spetsiaalne ensüüm, see kulutab ATP-d.
Saade(alates lat. saade- ülekanne) on polüpeptiidahela biosüntees mRNA maatriksil, mille käigus geneetiline informatsioon transleeritakse polüpeptiidahela aminohappejärjestuseks.

Valgu sünteesi teine etapp toimub kõige sagedamini tsütoplasmas, näiteks krobelisel endoplasmaatilisel retikulumil. Selle esinemine eeldab ribosoomide olemasolu, tRNA aktiveerimist, mille käigus seovad need vastavad aminohapped, Mg2+ ioonide olemasolu, aga ka optimaalseid keskkonnatingimusi (temperatuur, pH, rõhk jne).
Saate alustada algatus) sünteesiks valmis mRNA molekuli külge kinnitub väike ribosoomi alaühik ja seejärel komplementaarsuse põhimõtte kohaselt selekteeritakse aminohapet metioniini kandev tRNA esimesse koodonisse (AUG). Alles siis liitub ribosoomi suur subühik. Kokkupandud ribosoomi sees on kaks mRNA koodonit, millest esimene on juba hõivatud. Sellega külgneva koodoni külge kinnitub teine, samuti aminohapet kandev tRNA, misjärel tekib ensüümide abil aminohappejääkide vahel peptiidside. Ribosoom liigutab ühte mRNA koodonit; esimene aminohappest vabastatud tRNA-st naaseb järgmise aminohappe saamiseks tsütoplasmasse ja ülejäänud tRNA küljes ripub justkui tulevase polüpeptiidahela fragment. Järgmine tRNA liitub uue koodoniga, mis on ribosoomi sees, protsess kordub ja samm-sammult polüpeptiidahel pikeneb, s.t. pikenemine.
Valgu sünteesi lõpp lõpetamine) tekib niipea, kui mRNA molekulis, mis ei kodeeri aminohapet (stoppkoodon), kohtab spetsiifilist nukleotiidjärjestust. Pärast seda eraldatakse ribosoom, mRNA ja polüpeptiidahel ning äsja sünteesitud valk omandab sobiva struktuuri ja transporditakse raku sellesse ossa, kus see hakkab oma funktsioone täitma.
Translatsioon on väga energiamahukas protsess, kuna ühe ATP molekuli energia kulub ühe aminohappe kinnitamiseks tRNA-le ja veel mitut kasutatakse ribosoomi liigutamiseks mööda mRNA molekuli.
Teatud valgumolekulide sünteesi kiirendamiseks saab mRNA molekuli külge järjestikku kinnitada mitu ribosoomi, mis moodustavad ühtse struktuuri - polüsoom.
Rakk on elusolendi geneetiline üksus. Kromosoomid, nende struktuur (kuju ja suurus) ja funktsioonid. Kromosoomide arv ja nende liigiline püsivus. Somaatilised ja sugurakud. Raku elutsükkel: interfaas ja mitoos. Mitoos on somaatiliste rakkude jagunemine. Meioos. Mitoosi ja meioosi faasid. Sugurakkude areng taimedes ja loomades. Rakkude jagunemine on organismide kasvu, arengu ja paljunemise aluseks. Meioosi ja mitoosi roll
Rakk on elu geneetiline üksus
Hoolimata asjaolust, et nukleiinhapped on geneetilise teabe kandjad, on selle teabe rakendamine väljaspool rakku võimatu, mida on lihtne tõestada viiruste näitel. Need organismid, mis sisaldavad sageli ainult DNA-d või RNA-d, ei saa iseseisvalt paljuneda, selleks peavad nad kasutama raku pärilikku aparaati. Nad ei suuda isegi rakku tungida ilma raku enda abita, välja arvatud membraanitranspordi mehhanisme kasutades või rakukahjustuse tõttu. Enamik viirusi on ebastabiilsed, nad surevad pärast mõnetunnist vabas õhus viibimist. Seetõttu on rakk elavate inimeste geneetiline üksus, millel on minimaalne komplekt komponendid päriliku teabe säilitamiseks, muutmiseks ja rakendamiseks, samuti selle edastamiseks järglastele.
Suurem osa eukarüootse raku geneetilisest informatsioonist asub tuumas. Selle organisatsiooni tunnuseks on see, et erinevalt prokarüootse raku DNA-st ei ole eukarüootsed DNA molekulid suletud ja moodustavad valkudega kompleksseid komplekse - kromosoome.
Kromosoomid, nende struktuur (kuju ja suurus) ja funktsioonid
Kromosoom(kreeka keelest. kroomitud- värv, värv ja säga- keha) on raku tuuma struktuur, mis sisaldab geene ja kannab teatud pärilikku teavet keha tunnuste ja omaduste kohta.
Mõnikord nimetatakse prokarüootide ring-DNA molekule ka kromosoomideks. Kromosoomid on võimelised ise paljunema, neil on struktuurne ja funktsionaalne individuaalsus ning nad säilitavad selle mitme põlvkonna jooksul. Iga rakk kannab endas kogu keha pärilikku teavet, kuid ainult väike osa sellest töötab.
Kromosoomi aluseks on kaheahelaline DNA molekul, mis on pakitud valkudega. Eukarüootides interakteeruvad histooni ja mittehistooni valgud DNA-ga, prokarüootides histooni valgud puuduvad.
Kromosoomid on kõige paremini nähtavad valgusmikroskoobi all rakkude jagunemise ajal, kui need muutuvad tihendamise tulemusena vardakujulisteks kehadeks, mis on eraldatud esmase kitsendusega - tsentromeer - õlgadele. Kromosoomil võib olla ka sekundaarne kitsendus, mis teatud juhtudel eraldab nn satelliit. Kromosoomide otsad on nn telomeerid. Telomeerid takistavad kromosoomide otste kokkukleepumist ja tagavad nende kinnitumise tuumamembraanile mittejagunevas rakus. Jagunemise alguses on kromosoomid kahekordistunud ja koosnevad kahest tütarkromosoomist - kromatiidid kinnitatud tsentromeerile.

Kuju järgi eristatakse võrdse käega, ebavõrdse käega ja vardakujulisi kromosoome. Kromosoomide suurused on märkimisväärselt erinevad, kuid keskmise kromosoomi suurus on 5 $ × $ 1,4 µm.
Mõnel juhul sisaldavad kromosoomid arvukate DNA dubleerimise tulemusena sadu ja tuhandeid kromatiide: selliseid hiiglaslikke kromosoome nimetatakse nn. polüetüleen. Neid leidub Drosophila vastsete süljenäärmetes, aga ka ümarusside seedenäärmetes.
Kromosoomide arv ja nende liigiline püsivus. Somaatilised ja sugurakud
Rakuteooria järgi on rakk organismi struktuuri, elu ja arengu üksus. Seega on rakutasandil tagatud elusolendite sellised olulised funktsioonid nagu organismi kasv, paljunemine ja areng. Rakud mitmerakulised organismid võib jagada somaatiliseks ja seksuaalseks.
somaatilised rakud on kõik keharakud, mis tekivad mitootilise jagunemise tulemusena.
Kromosoomide uurimine võimaldas kindlaks teha, et iga bioloogilise liigi organismi somaatilisi rakke iseloomustab konstantne kromosoomide arv. Näiteks inimesel on neid 46. Somaatiliste rakkude kromosoomide komplekti nimetatakse diploidne(2n) või topelt.

sugurakud, või sugurakud, on spetsiaalsed rakud, mis teenivad seksuaalset paljunemist.
Sugurakud sisaldavad alati poole vähem kromosoome kui somaatilistes rakkudes (inimesel - 23), seega nimetatakse sugurakkude kromosoomide komplekti nn. haploidne(n) või üksik. Selle teket seostatakse meiootiliste rakkude jagunemisega.
Somaatiliste rakkude DNA kogus on tähistatud kui 2c ja sugurakkude DNA kogus 1c. Somaatiliste rakkude geneetiline valem on kirjutatud kui 2n2c ja sugu - 1n1c.
Mõnede somaatiliste rakkude tuumades võib kromosoomide arv erineda nende arvust somaatilistes rakkudes. Kui see erinevus on ühe, kahe, kolme jne haploidsete hulga võrra suurem, siis nimetatakse selliseid rakke polüploidne(vastavalt tri-, tetra-, pentaploidne). Sellistes rakkudes on ainevahetusprotsessid tavaliselt väga intensiivsed.
Kromosoomide arv iseenesest ei ole liigispetsiifiline tunnus, kuna erinevatel organismidel võib olla võrdne arv kromosoome, samas kui lähedastel organismidel võib olla erinev arv. Näiteks malaariaplasmoodiumil ja hobuste ümarussil on kaks kromosoomi, inimestel ja šimpansil vastavalt 46 ja 48.
Inimese kromosoomid jagunevad kahte rühma: autosoomid ja sugukromosoomid (heterokromosoomid). Autosoom inimese somaatilistes rakkudes on 22 paari, need on meestel ja naistel ühesugused ning sugukromosoomid ainult üks paar, kuid tema määrab isendi soo. Sugukromosoome on kahte tüüpi - X ja Y. Naise keharakud kannavad kahte X-kromosoomi ja meestel X- ja Y-kromosoomi.
Karüotüüp- see on organismi kromosoomikomplekti märkide kogum (kromosoomide arv, kuju ja suurus).
Kariotüübi tingimuslik rekord sisaldab kromosoomide koguarvu, sugukromosoome ja võimalikke kõrvalekaldeid kromosoomide komplektis. Näiteks normaalse mehe karüotüüp on kirjutatud 46, XY ja karüotüüp normaalne naine- 46, XX.
Raku elutsükkel: interfaas ja mitoos
Rakud ei teki iga kord uuesti, need tekivad ainult emarakkude jagunemise tulemusena. Pärast eraldamist kulub tütarrakkudel veidi aega, et moodustada organellid ja omandada sobiv struktuur, mis tagaks teatud funktsiooni täitmise. Seda ajaperioodi nimetatakse valmimine.
Nimetatakse ajavahemikku raku ilmumisest jagunemise tulemusena kuni selle jagunemiseni või surmani raku elutsükkel.
Eukarüootsetes rakkudes jaguneb elutsükkel kaheks põhifaasiks: interfaas ja mitoos.
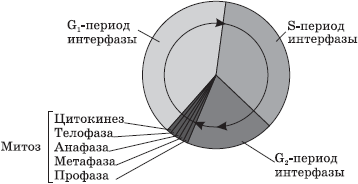
Interfaas- see on ajavahemik elutsüklis, mille jooksul rakk ei jagune ja toimib normaalselt. Interfaas jaguneb kolmeks perioodiks: G 1 -, S- ja G 2 -periood.
G 1 -periood(presünteetiline, postmitootiline) on rakkude kasvu ja arengu periood, mille jooksul toimub aktiivne RNA, valkude ja muude ainete süntees, mis on vajalikud vastloodud raku täielikuks elutegevuseks. Selle perioodi lõpuks võib rakk hakata valmistuma DNA dubleerimiseks.
AT S-periood(sünteetiline) toimub DNA replikatsiooni protsess. Ainus kromosoomi osa, mis ei replikatsiooni ei toimu, on tsentromeer, mistõttu tekkivad DNA molekulid ei lahkne täielikult, vaid jäävad sellesse kinni ning jagunemise alguses on kromosoom X-kujuline. Raku geneetiline valem pärast DNA dubleerimist on 2n4c. Ka S-perioodil toimub rakukeskuse tsentrioolide kahekordistumine.
G 2 -periood(postsünteetiline, premitootiline) iseloomustab raku jagunemise protsessiks vajalik RNA, valkude ja ATP intensiivne süntees, samuti tsentrioolide, mitokondrite ja plastiidide eraldamine. Kuni interfaasi lõpuni jäävad kromatiin ja nukleool selgelt eristatavaks, tuumamembraani terviklikkus ei ole häiritud ja organellid ei muutu.
Mõned keharakud on võimelised täitma oma funktsioone kogu keha eluea jooksul (meie aju neuronid, südame lihasrakud), teised eksisteerivad lühikest aega, misjärel surevad (sooleepiteeli rakud). , naha epidermise rakud). Järelikult peavad organismis pidevalt toimuma rakkude jagunemise protsessid ja uute rakkude teke, mis surnud asendaksid. Jagunemisvõimelisi rakke nimetatakse varre. Inimese kehas leidub neid punases luuüdis, naha epidermise sügavates kihtides ja mujal. Neid rakke kasutades saate kasvatada uue organi, saavutada noorendamise ja ka keha kloonida. Tüvirakkude kasutamise väljavaated on üsna selged, kuid selle probleemi moraalseid ja eetilisi aspekte arutatakse endiselt, kuna enamasti kasutatakse abordi käigus hukkunud inimese embrüotest saadud embrüonaalseid tüvirakke.
Interfaasi kestus taime- ja loomarakkudes on keskmiselt 10–20 tundi, mitoos aga umbes 1–2 tundi.
Mitmerakuliste organismide järjestikuste jagunemiste käigus muutuvad tütarrakud üha mitmekesisemaks, kuna nad loevad teavet kõigest. rohkem geenid.
Mõned rakud lõpetavad aja jooksul jagunemise ja surevad, mis võib olla tingitud teatud funktsioonide täitumisest, näiteks naha epidermaalsete rakkude ja vererakkude puhul, või nende rakkude kahjustamisest tegurite mõjul. keskkond, eriti patogeenid. Geneetiliselt programmeeritud rakusurma nimetatakse apoptoos, kui juhuslik surm - nekroos.
Mitoos on somaatiliste rakkude jagunemine. Mitoosi faasid
Mitoos- somaatiliste rakkude kaudse jagunemise meetod.
Mitoosi käigus läbib rakk järjestikuste faaside jada, mille tulemusena saab iga tütarrakk samasuguse kromosoomikomplekti, mis emarakus.
Mitoos jaguneb neljaks põhifaasiks: profaas, metafaas, anafaas ja telofaas. Profaas- pikim mitoosi staadium, mille käigus toimub kromatiini kondenseerumine, mille tulemusena tulevad nähtavale kahest kromatiidist (tütarkromosoomidest) koosnevad X-kujulised kromosoomid. Sel juhul tuum kaob, tsentrioolid lahknevad raku pooluste suunas ja hakkab moodustuma mikrotuubulite akromatiini spindel (spindel). Profaasi lõpus laguneb tuumamembraan eraldi vesiikuliteks.

AT metafaas kromosoomid joonduvad piki raku ekvaatorit oma tsentromeeridega, mille külge kinnituvad täielikult moodustunud jagunemisspindli mikrotuubulid. Selles jagunemisetapis on kromosoomid kõige tihedamad ja iseloomuliku kujuga, mis võimaldab uurida karüotüüpi.
AT anafaasis tsentromeerides toimub kiire DNA replikatsioon, mille tulemusena kromosoomid lõhenevad ja kromatiidid lahknevad mikrotuubulitega venitatuna raku pooluste suunas. Kromatiidide jaotus peab olema absoluutselt võrdne, kuna just see protsess säilitab kromosoomide arvu püsivuse keharakkudes.
Laval telofaas tütarkromosoomid kogunevad poolustele, despiraliseeruvad, nende ümber moodustuvad vesiikulitest tuumaümbrised ja äsja moodustunud tuumadesse tekivad tuumakesed.
Pärast tuuma jagunemist toimub tsütoplasma jagunemine - tsütokinees, mille käigus toimub emaraku kõigi organellide enam-vähem ühtlane jaotus.
Seega moodustub mitoosi tulemusena ühest emarakust kaks tütarrakku, millest igaüks on emaraku geneetiline koopia (2n2c).
Haigetes, kahjustatud, vananevates rakkudes ja keha spetsialiseeritud kudedes võib toimuda veidi erinev jagunemisprotsess - amitoos. Amitoos nimetatakse eukarüootsete rakkude otseseks jagunemiseks, kus geneetiliselt ekvivalentsete rakkude moodustumist ei toimu, kuna rakukomponendid jaotuvad ebaühtlaselt. See esineb taimedes endospermis ja loomadel maksas, kõhres ja silma sarvkestas.
Meioos. Meioosi faasid
Meioos- see on primaarsete sugurakkude (2n2c) kaudse jagunemise meetod, mille tulemusena moodustuvad haploidsed rakud (1n1c), kõige sagedamini sugurakud.
Erinevalt mitoosist koosneb meioos kahest järjestikusest raku jagunemisest, millest igaühele eelneb interfaas. Meioosi esimest jagunemist (meioosi I) nimetatakse vähendamine, kuna sel juhul väheneb kromosoomide arv poole võrra ja teine jagunemine (meioos II) - võrrand, kuna selle käigus säilib kromosoomide arv.

I interfaas kulgeb sarnaselt mitoosi interfaasiga. Meioos I jaguneb neljaks faasiks: profaas I, metafaas I, anafaas I ja telofaas I. profaas I toimub kaks peamist protsessi – konjugatsioon ja üleminek. Konjugatsioon- see on homoloogsete (paaritud) kromosoomide liitmise protsess kogu pikkuses. Konjugatsiooni käigus tekkinud kromosoomipaarid säilivad kuni metafaasi I lõpuni.
Üleminek- homoloogsete kromosoomide homoloogsete piirkondade vastastikune vahetus. Ülekandmise tulemusena omandavad mõlemalt vanemalt organismi saadud kromosoomid uued geenikombinatsioonid, mis toob kaasa geneetiliselt mitmekesiste järglaste ilmumise. Profaasi I lõpus, nagu ka mitoosi profaasis, kaob tuum, tsentrioolid lahknevad raku pooluste suunas ja tuumaümbris laguneb.

AT metafaas I kromosoomipaarid reastuvad piki raku ekvaatorit, nende tsentromeeride külge on kinnitatud spindli mikrotuubulid.
AT anafaas I kahest kromatiidist koosnevad terved homoloogsed kromosoomid lahknevad poolustele.
AT telofaas I kromosoomide klastrite ümber raku poolustes tekivad tuumamembraanid, moodustuvad tuumakesed.
Tsütokinees I tagab tütarrakkude tsütoplasmade jagunemise.
Meioosi I (1n2c) tulemusena tekkinud tütarrakud on geneetiliselt heterogeensed, kuna nende kromosoomid, mis on juhuslikult raku poolustele hajutatud, sisaldavad ebavõrdseid geene.
Mitoosi ja meioosi võrdlevad omadused
| märk | Mitoos | Meioos | |
| Millised rakud hakkavad jagunema? | Somaatiline (2n) | Primaarsed sugurakud (2n) | |
| Osakondade arv | 1 | 2 | |
| Kui palju ja milliseid rakke tekib jagunemise käigus? | 2 somaatilist (2n) | 4 seksuaalset (n) | |
| Interfaas | Rakkude ettevalmistamine jagunemiseks, DNA dubleerimiseks | Väga lühike, DNA dubleerimist ei toimu | |
| Faasid | Meioos I | Meioos II | |
| Profaas | Võib tekkida kromosoomide kondenseerumine, tuuma kadumine, tuumaümbrise lagunemine, konjugatsioon ja ristumine | Kromosoomide kondenseerumine, tuuma kadumine, tuumaümbrise lagunemine | |
| metafaas | Kromosoomipaarid paiknevad piki ekvaatorit, moodustub jaotusvõll | Kromosoomid reastuvad piki ekvaatorit, moodustub jagunemisvõll | |
| Anafaas | Kahe kromatiidi homoloogsed kromosoomid lahknevad pooluste suunas | Kromatiidid lahknevad pooluste suunas | |
| Telofaas | Kromosoomid despiraliseerivad, moodustuvad uued tuumaümbrised ja nukleoolid | Kromosoomid despiraliseerivad, moodustuvad uued tuumaümbrised ja nukleoolid | |
II faas väga lühike, kuna selles DNA kahekordistumist ei toimu, st S-perioodi pole.
Meioos II samuti jagatud neljaks faasiks: II faas, II metafaas, II anafaas ja II telofaas. AT profaas II toimuvad samad protsessid, mis I profaasis, välja arvatud konjugatsioon ja üleminek.
AT metafaas II Kromosoomid asuvad piki raku ekvaatorit.
AT anafaas II Kromosoomid lõhenevad tsentromeeril ja kromatiidid venivad pooluste suunas.
AT telofaas II Tütarkromosoomide klastrite ümber moodustuvad tuumamembraanid ja nukleoolid.
Pärast tsütokinees II kõigi nelja tütarraku geneetiline valem on 1n1c, kuid neil kõigil on erinev geenide komplekt, mis tuleneb ema- ja isakromosoomide ristumisest ja juhuslikust kombinatsioonist tütarrakkudes.
Sugurakkude areng taimedes ja loomades
Gametogenees(kreeka keelest. sugurakud- naine, sugurakud- abikaasa ja genees- päritolu, esinemine) on küpsete sugurakkude moodustumise protsess.
Kuna sugulisel paljunemisel on enamasti vaja kahte isendit - emast ja isast, kes toodavad erinevaid sugurakke - munarakke ja spermat, siis peaksid nende sugurakkude moodustumise protsessid olema erinevad.
Protsessi iseloom sõltub suuresti sellest, kas see toimub taimes või looma puur, kuna taimedes toimub sugurakkude moodustumise ajal ainult mitoos ja loomadel - nii mitoos kui ka meioos.
Sugurakkude areng taimedes. Kaasseemnetaimedes moodustuvad isas- ja emased sugurakud aastal erinevad osad lill - vastavalt tolmukad ja püstolid.
Enne meeste sugurakkude moodustumist - mikrogametogenees(kreeka keelest. mikros- väike) - toimub mikrosporogenees, see tähendab tolmukate tolmukatesse mikroeoste teket. Seda protsessi seostatakse emaraku meiootilise jagunemisega, mille tulemuseks on neli haploidset mikrospoori. Mikrogametogenees on seotud mikrospooride mitootilise jagunemisega, mis annab kahest suurest rakust koosneva isase gametofüüdi. vegetatiivne(sifonogeenne) ja madal generatiivne. Isasgametofüüt on pärast jagunemist kaetud tihedate kestadega ja moodustab õietolmutera. Mõnel juhul jaguneb generatiivne rakk mitootiliselt isegi õietolmu küpsemise protsessis ja mõnikord alles pärast emaka häbimärgisesse ülekandmist, moodustades kaks liikumatut isase sugurakku - sperma. Pärast tolmeldamist moodustub vegetatiivsest rakust õietolmutoru, mille kaudu tungivad spermatosoidid viljastamiseks pesa munasarja.

Naiste sugurakkude arengut taimedes nimetatakse megagametogenees(kreeka keelest. megas- suur). See esineb pesa munasarjas, millele eelneb megasporogenees, mille tulemusena moodustub tuumas paikneva megaspoori emarakust meiootilise jagunemise teel neli megaspoori. Üks megaspooridest jaguneb mitootiliselt kolm korda, andes emasele gametofüüdile kaheksa tuumaga embrüokoti. Järgneva tütarrakkude tsütoplasmade isoleerimisega saab ühest saadud rakkudest munarakk, mille külgedel asuvad nn sünergiidid, embrüokoti vastasotsas moodustuvad kolm antipoodi ja keskel. , kahe haploidse tuuma ühinemise tulemusena moodustub diploidne keskrakk.

Sugurakkude areng loomadel. Loomadel eristatakse kahte sugurakkude moodustumise protsessi - spermatogeneesi ja oogeneesi.
spermatogenees(kreeka keelest. sperma, spermatosoidid- seeme ja genees- päritolu, esinemine) on küpsete meessoost sugurakkude - spermatosoidide moodustumise protsess. Inimestel esineb see munandites ehk munandites ning jaguneb neljaks perioodiks: paljunemine, kasv, küpsemine ja moodustumine.

AT paaritumis hooaegürgsed sugurakud jagunevad mitootiliselt, mille tulemusena moodustuvad diploidid spermatogoonia. AT kasvuperiood spermatogooniad koguvad toitaineid tsütoplasmasse, suurenevad ja muutuvad primaarsed spermatotsüüdid, või I järgu spermatotsüüdid. Alles pärast seda sisenevad nad meioosi ( valmimisperiood), mille tulemuseks on esmalt kaks sekundaarne spermatotsüüt, või 2. järku spermatotsüüdid ja seejärel - neli haploidset rakku, millel on üsna palju tsütoplasma - spermatiidid. AT kujunemisperiood nad kaotavad peaaegu kogu tsütoplasma ja moodustavad lipu, mis muutub spermatosoidideks.
spermatosoidid, või kummikommid, - väga väikesed liikuvad isassugurakud pea, kaela ja sabaga.

AT pea, välja arvatud tuum, on akrosoom- modifitseeritud Golgi kompleks, mis tagab munaraku membraanide lahustumise viljastamise ajal. AT kaela seal on rakukeskuse tsentrioolid ja alus hobusesaba moodustavad mikrotuubulid, mis toetavad otseselt spermatosoidide liikumist. See sisaldab ka mitokondreid, mis annavad sperma liikumiseks ATP-energiat.
Ovogenees(kreeka keelest. ÜRO- muna ja genees- päritolu, esinemine) on küpsete naiste sugurakkude - munade moodustumise protsess. Inimestel esineb see munasarjades ja koosneb kolmest perioodist: paljunemine, kasv ja küpsemine. Spermatogeneesiga sarnased paljunemis- ja kasvuperioodid esinevad isegi emakasisese arengu ajal. Samal ajal tekivad mitoosi tulemusena esmastest sugurakkudest diploidsed rakud. oogonia, mis seejärel muutuvad diploidseks primaarseks munarakud, või 1. järku munarakud. aastal esinev meioos ja sellele järgnev tsütokinees valmimisperiood, mida iseloomustab emaraku tsütoplasma ebaühtlane jagunemine, nii et selle tulemusel saadakse algul üks sekundaarne munarakk, või munarakk 2. järk ja esimene polaarkeha, ja seejärel sekundaarsest munarakust - munarakk, mis säilitab kogu varu toitaineid, ja teine polaarkeha, samas kui esimene polaarkeha on jagatud kaheks. Polaarkehad võtavad ära liigse geneetilise materjali.
Inimestel tekivad munad 28–29-päevase intervalliga. Munarakkude küpsemise ja vabanemisega seotud tsüklit nimetatakse menstruaaltsükliks.
Muna- suur naissoost sugurakk, mis ei kanna mitte ainult haploidset kromosoomide komplekti, vaid ka olulist toitainete pakkumist embrüo edasiseks arenguks.

Imetajate muna on kaetud nelja membraaniga, mis vähendab erinevate tegurite mõjul selle kahjustamise tõenäosust. Muna läbimõõt inimestel ulatub 150–200 mikronini, jaanalinnul võib see olla mitu sentimeetrit.
Rakkude jagunemine on organismide kasvu, arengu ja paljunemise aluseks. Mitoosi ja meioosi roll
Kui üherakulistes organismides põhjustab rakkude jagunemine isendite arvu suurenemist, s.t paljunemist, siis hulkraksetes organismides võib see protsess olla erinev tähendus. Seega on embrüo rakkude jagunemine, alustades sügoodist, bioloogiline alus omavahel seotud kasvu- ja arenguprotsessidele. Sarnaseid muutusi täheldatakse inimesel noorukieas, kui mitte ainult ei suurene rakkude arv, vaid toimub ka kvalitatiivne muutus kehas. Rakkude jagunemise aluseks on ka hulkraksete organismide paljunemine, näiteks mittesugulisel paljunemisel taastub selle protsessi tõttu mingist kehaosast terve keha ning sugulisel paljunemisel tekivad gametogeneesi käigus sugurakud, mis järgnevalt annavad. uus organism. Tuleb märkida, et eukarüootsete rakkude jagunemise peamised meetodid - mitoos ja meioos - on erineva tähendusega. elutsüklid organismid.
Mitoosi tulemusena toimub päriliku materjali ühtlane jaotumine tütarrakkude vahel - täpsed koopiad emalik. Ilma mitoosita oleks ühest rakust - sigootist - arenevate mitmerakuliste organismide olemasolu ja kasv võimatu, kuna kõik selliste organismide rakud peavad sisaldama sama geneetilist teavet.
Jagunemise käigus muutuvad tütarrakud struktuurilt ja funktsioonidelt järjest mitmekesisemaks, mis on seotud rakkudevahelise interaktsiooni tõttu neis uute geenirühmade aktiveerumisega. Seega on mitoos organismi arenguks vajalik.
See rakkude jagunemise meetod on protsesside jaoks vajalik mittesuguline paljunemine ning kahjustatud kudede ja elundite taastamine (taastamine).
Meioos omakorda tagab karüotüübi püsivuse sugulisel paljunemisel, kuna vähendab poole võrra enne sugulist paljunemist kromosoomide komplekti, mis seejärel viljastamise tulemusena taastub. Lisaks põhjustab meioos vanemate geenide uute kombinatsioonide ilmnemist, mis on tingitud kromosoomide ristumisest ja juhuslikust kombinatsioonist tütarrakkudes. Tänu sellele on järglased geneetiliselt mitmekesised, mis annab materjali looduslik valik ja see on evolutsiooni materiaalne alus. Kromosoomide arvu, kuju ja suuruse muutumine võib ühelt poolt kaasa tuua erinevate kõrvalekallete ilmnemise organismi arengus ja isegi selle surma, teisalt aga indiviidide ilmumiseni. keskkonnaga paremini kohanenud.
Seega on rakk organismide kasvu, arengu ja paljunemise üksus.
Iga valku esindab üks või mitu polüpeptiidahelat. DNA osa, mis kannab teavet ühe polüpeptiidahela kohta, nimetatakse genoom. DNA molekulide kogum rakus toimib geneetilise informatsiooni kandjana. Geneetiline teave edastatakse emarakkudelt tütarrakkudele ja vanematelt lastele. Geen on geneetika ühik, või pärilikku teavet.
DNA on geneetilise informatsiooni kandja rakus- ei osale otseselt valkude sünteesis. Eukarüootsetes rakkudes sisalduvad DNA molekulid tuuma kromosoomides ja eraldatakse tuumamembraaniga tsütoplasmast, kus sünteesitakse valke. Ribosoomidesse - valkude kogunemiskohtadesse - saadetakse tuumast infokandja, mis on võimeline läbima tuumaümbrise poore. Messenger RNA (mRNA) on selline vahendaja. Komplementaarsuse põhimõtte kohaselt sünteesitakse see DNA-l ensüümi RNA- osalusel. polümeraas.
1) RNA sünteesi protsessi, mille käigus ühte DNA molekuli ahelatest kasutatakse mallina, nimetatakse nn. transkriptsioon .
2) Transkriptsioon - see on mehhanism, mille abil ühe DNA ahela nukleotiidjärjestus kirjutatakse ümber mRNA molekuli komplementaarse järjestuse poolt.
Messenger RNA on üheahelaline molekul ja transkriptsioon pärineb kaheahelalise DNA molekuli ühest ahelast. See ei ole terve DNA molekuli koopia, vaid ainult osa sellest - üks geen eukarüootides või külgnevate geenide rühm, mis kannab teavet valkude struktuuri kohta, mis on vajalikud ühe funktsiooni täitmiseks prokarüootides. Seda geenide rühma nimetatakse operon. Iga operoni alguses on omamoodi RNA polümeraasi maandumiskoht, mida nimetatakse promootor.see on spetsiifiline DNA nukleotiidide järjestus, mille ensüüm "ära tunneb" keemilise afiinsuse tõttu. Ainult promootori külge kinnitumisel on RNA polümeraas võimeline käivitama RNA sünteesi. Olles jõudnud operoni lõppu, kohtab ensüüm signaali (teatud nukleotiidide järjestuse kujul), mis näitab lugemise lõppu. Valmis mRNA eemaldub DNA-st ja läheb valgusünteesi kohta.
Transkriptsiooniprotsessis on neli etappi: 1) RNA sidumine- polümeraas koos promootoriga; 2) algatus- sünteesi algus. See seisneb esimese fosfodiestersideme moodustamises ATP või GTP ja sünteesitud RNA molekuli teise nukleotiidi vahel; 3) pikenemine– RNA ahela kasv; need. nukleotiidide järjestikune lisamine üksteisele selles järjekorras, milles nende komplementaarsed nukleotiidid on transkribeeritud DNA ahelas. Pikenduskiirus on 50 nukleotiidi sekundis; neli) lõpetamine- RNA sünteesi lõpetamine.
Pärast tuumaümbrise pooride läbimist saadetakse mRNA ribosoomidesse, kus dešifreeritakse geneetiline informatsioon - see tõlgitakse nukleotiidide "keelest" aminohapete "keelde". Ribosoomides esinev polüpeptiidahelate süntees mRNA matriitsi järgi on nn. saade(lat. tõlge - tõlge).
Aminohapped, millest sünteesitakse valke, viiakse ribosoomidesse spetsiaalsete RNA-de abil, mida nimetatakse transport-RNA-deks (tRNA). Erinevaid tRNA-sid on rakus sama palju kui aminohappeid kodeerivaid koodoneid. Iga tRNA "lehe" ülaosas on kolmest nukleotiidist koosnev järjestus, mis on komplementaarsed mRNA koodoni nukleotiididega. Nad kutsuvad teda antikoodon. Spetsiaalne ensüüm – kodaas – tunneb ära tRNA ja kinnitab "lehelehele" aminohappe – ainult selle, mida kodeerib antikoodoniga komplementaarne kolmik. Ühe ATP molekuli energia kulutatakse kovalentse sideme moodustamiseks tRNA ja selle "oma" aminohappe vahel.
 Selleks, et aminohape sisalduks polüpeptiidahelas, peab see tRNA-st lahku lööma. See saab võimalikuks, kui tRNA siseneb ribosoomi ja antikoodon tunneb mRNA-s ära oma koodoni. Ribosoomil on kaks kohta kahe tRNA molekuli sidumiseks. Üks neist valdkondadest, nn aktsepteerija tRNA siseneb koos aminohappega ja kinnitub selle koodoniga (I). Kas see aminohape kinnitub enda külge (aktsepteerib) kasvavat valguahelat (II)? Nende vahel moodustub peptiidside. tRNA, mis on nüüd ühendatud koos mRNA koodoniga doonor ribosoomi osa. Vabanenud aktseptorsaiti tuleb uus tRNA, mis on seotud aminohappega, mille krüpteerib järgmine koodon (III). Doonorikohast kantakse eraldunud polüpeptiidahel uuesti üle siia ja pikendatakse veel ühe lüli võrra. Kasvavas ahelas olevad aminohapped on ühendatud järjestuses, milles neid kodeerivad koodonid asuvad mRNA-s.
Selleks, et aminohape sisalduks polüpeptiidahelas, peab see tRNA-st lahku lööma. See saab võimalikuks, kui tRNA siseneb ribosoomi ja antikoodon tunneb mRNA-s ära oma koodoni. Ribosoomil on kaks kohta kahe tRNA molekuli sidumiseks. Üks neist valdkondadest, nn aktsepteerija tRNA siseneb koos aminohappega ja kinnitub selle koodoniga (I). Kas see aminohape kinnitub enda külge (aktsepteerib) kasvavat valguahelat (II)? Nende vahel moodustub peptiidside. tRNA, mis on nüüd ühendatud koos mRNA koodoniga doonor ribosoomi osa. Vabanenud aktseptorsaiti tuleb uus tRNA, mis on seotud aminohappega, mille krüpteerib järgmine koodon (III). Doonorikohast kantakse eraldunud polüpeptiidahel uuesti üle siia ja pikendatakse veel ühe lüli võrra. Kasvavas ahelas olevad aminohapped on ühendatud järjestuses, milles neid kodeerivad koodonid asuvad mRNA-s.
Kui ribosoomilt leitakse üks kolmest kolmikust ( UAA, UAG, UGA), mis on geenidevahelised "kirjavahemärgid", ei saa ükski tRNA aktseptorsaidis asuda. Fakt on see, et puuduvad antikoodonid, mis oleksid komplementaarsed "kirjavahemärkide" nukleotiidjärjestustega. Eraldunud ahelal pole aktseptorkohas millegagi kinnituda ja see lahkub ribosoomist. Valkude süntees on lõppenud.
Prokarüootides algab valgusüntees koodonist AUG, mis asub iga geeni koopias esimesel kohal, on ribosoomis sellisel positsioonil, et spetsiaalse tRNA antikoodon interakteerub sellega, mis on seotud formüülmentioniin. See aminohappe metioniini modifitseeritud vorm siseneb koheselt doonorikohta ja mängib fraasis suure algustähe rolli - mis tahes polüpeptiidahela süntees algab sellega bakterirakus. Kui kolmik AUG ei ole esiteks, vaid geeni koopia sees kodeerib see aminohapet metioniini. Pärast polüpeptiidahela sünteesi lõppu eraldatakse formüülmetioniin sellest ja seda valmisvalgus ei leidu.
 Valkude tootmise suurendamiseks läbib mRNA sageli korraga mitte ühte, vaid mitut ribosoomi. Millist struktuuri, mida ühendab üks mRNA molekul, nimetatakse polüsoom. Igal ribosoomil sünteesitakse sellel helmelaadsel konveieril identsed valgud.
Valkude tootmise suurendamiseks läbib mRNA sageli korraga mitte ühte, vaid mitut ribosoomi. Millist struktuuri, mida ühendab üks mRNA molekul, nimetatakse polüsoom. Igal ribosoomil sünteesitakse sellel helmelaadsel konveieril identsed valgud.
Aminohapped tarnitakse ribosoomidesse pidevalt tRNA kaudu. Pärast aminohappe annetamist lahkub tRNA ribosoomist ja ühendatakse kodaasi abil. Kõigi valkude tootmiseks vajalike "taimeteenuste" kõrge sidusus võimaldab mõne sekundi jooksul sünteesida sadadest aminohapetest koosnevaid polüpeptiidahelaid.
Geneetilise koodi omadused. Transkriptsiooniprotsessi kaudu rakus kantakse teave DNA-st üle valku.
DNA → mRNA → valk
DNA-s ja mRNA-s sisalduv geneetiline informatsioon sisaldub molekulide nukleotiidide järjestuses.
 Kuidas toimub informatsiooni tõlkimine nukleotiidide "keelest" aminohapete "keelde"? See tõlge viiakse läbi geneetilise koodi abil. kood või šifr, on sümbolite süsteem ühe teabevormi teiseks tõlkimiseks. Geneetiline kood on süsteem teabe salvestamiseks valkude aminohapete järjestuse kohta, kasutades mRNA nukleotiidide järjestust.
Kuidas toimub informatsiooni tõlkimine nukleotiidide "keelest" aminohapete "keelde"? See tõlge viiakse läbi geneetilise koodi abil. kood või šifr, on sümbolite süsteem ühe teabevormi teiseks tõlkimiseks. Geneetiline kood on süsteem teabe salvestamiseks valkude aminohapete järjestuse kohta, kasutades mRNA nukleotiidide järjestust.
Millised on geneetilise koodi omadused?
1. kolmiku kood. RNA sisaldab nelja nukleotiidi: A, G, C, W. Kui prooviksime määrata ühte aminohapet ühe nukleotiidiga, jääks 16 aminohapet 20-st krüpteerimata. Kahetäheline kood krüpteerib 16 aminohapet. Loodus on loonud kolmetähelise ehk kolmiku koodi. See tähendab et iga 20 aminohappest on kodeeritud kolmest nukleotiidist koosneva järjestusega, mida nimetatakse tripletiks või koodoniks.
2. Kood on degenereerunud. See tähendab et iga aminohapet kodeerib rohkem kui üks koodon. Erandid: meteoniin ja trüptofaan, millest igaüks on kodeeritud ühe kolmikuga.
3. Kood on üheselt mõistetav. Iga koodon kodeerib ainult ühte aminohapet.
4. Geenide vahel on "kirjavahemärgid". Trükitekstis on iga fraasi lõpus punkt. Mitmed seotud fraasid moodustavad lõigu. Geneetilise informatsiooni keeles on selline lõik operon ja seda komplementaarne mRNA. Iga geen prokarüootses operonis või üksikus eukarüootses geenis kodeerib ühte polüpeptiidahelat – fraasi. Kuna mõnel juhul luuakse mRNA matriitsile järjestikku mitu erinevat polüpeptiidahelat, tuleb need üksteisest eraldada. Selleks on geneetilisel aastal kolm erilist kolmikut - UAA, UAG, UGA, millest igaüks näitab ühe polüpeptiidahela sünteesi katkemist. Seega täidavad need kolmikud kirjavahemärkide funktsiooni. Need on iga geeni lõpus.
5. Geeni sees pole "vahemärke".
6. Kood on universaalne. Geneetiline kood on kõigil Maal elavatel olenditel sama. Bakterites ja seentes, nisus ja puuvillas, kalades ja ussides, konnades ja inimestes kodeerivad samad kolmikud samu aminohappeid.
 DNA replikatsiooni põhimõtted.
Protsess tagab geneetilise materjali järjepidevuse rakkude ja organismide põlvkondades replikatsioon – DNA molekulide dubleerimine. Seda keerukat protsessi viib läbi mitme ensüümi ja valgu kompleks, millel puudub katalüütiline aktiivsus ja mis on vajalikud polünukleotiidahelatele soovitud konformatsiooni saamiseks. Replikatsiooni tulemusena moodustuvad kaks identset DNA kaksikheeliksit. Need niinimetatud tütarmolekulid ei erine üksteisest ega algsest DNA algmolekulist. Replikatsioon toimub rakus enne jagunemist, seega saab iga tütarrakk täpselt samad DNA molekulid, mis emarakul. Replikatsiooniprotsess põhineb mitmel põhimõttel:
DNA replikatsiooni põhimõtted.
Protsess tagab geneetilise materjali järjepidevuse rakkude ja organismide põlvkondades replikatsioon – DNA molekulide dubleerimine. Seda keerukat protsessi viib läbi mitme ensüümi ja valgu kompleks, millel puudub katalüütiline aktiivsus ja mis on vajalikud polünukleotiidahelatele soovitud konformatsiooni saamiseks. Replikatsiooni tulemusena moodustuvad kaks identset DNA kaksikheeliksit. Need niinimetatud tütarmolekulid ei erine üksteisest ega algsest DNA algmolekulist. Replikatsioon toimub rakus enne jagunemist, seega saab iga tütarrakk täpselt samad DNA molekulid, mis emarakul. Replikatsiooniprotsess põhineb mitmel põhimõttel:
1. täiendavus. DNA lähtemolekuli mõlemad ahelad toimivad mallina selle komplementaarse, st. täiendav, tütarkett.
2. poolkonservatiivne. Replikatsiooni tulemusena moodustuvad kaks tütar-kaksikahelat, millest igaüks säilitab (säilitab) ühe algse DNA ahela muutumatuna. Tütarmolekulide teised ahelad sünteesitakse uuesti nukleotiididest mööda komplementaarsust algse DNA ahelaga. Tütar-DNA ei erine üksteisest ega algse kaksikheeliksist.
3.
 antiparallelism. Igal DNA ahelal on spetsiifiline orientatsioon. Üks ots kannab hüdroksüülrühma (-OH), mis on seotud suhkru desoksüriboosi 3'-süsinikuga, ahela teises otsas on suhkru 5'-asendis fosforhappejääk. Kaks komplementaarset ahelat DNA molekulis on orienteeritud vastassuunas – antiparalleelselt. Ensüümid, mis sünteesivad uusi DNA ahelaid ja mida nimetatakse DNA polümeraasideks, võivad liikuda piki matriitsi ahelaid ainult ühes suunas – nende 3'-otstest 5'-otsteni. Sel juhul toimub komplementaarsete ahelate süntees alati 5'→3' suunas, s.o. unipolaarne. Seetõttu toimub replikatsiooniprotsessis uute ahelate samaaegne süntees antiparalleelselt.
antiparallelism. Igal DNA ahelal on spetsiifiline orientatsioon. Üks ots kannab hüdroksüülrühma (-OH), mis on seotud suhkru desoksüriboosi 3'-süsinikuga, ahela teises otsas on suhkru 5'-asendis fosforhappejääk. Kaks komplementaarset ahelat DNA molekulis on orienteeritud vastassuunas – antiparalleelselt. Ensüümid, mis sünteesivad uusi DNA ahelaid ja mida nimetatakse DNA polümeraasideks, võivad liikuda piki matriitsi ahelaid ainult ühes suunas – nende 3'-otstest 5'-otsteni. Sel juhul toimub komplementaarsete ahelate süntees alati 5'→3' suunas, s.o. unipolaarne. Seetõttu toimub replikatsiooniprotsessis uute ahelate samaaegne süntees antiparalleelselt.
4. katkestus. Selleks, et uued DNA ahelad saaksid üles ehitada komplementaarsuse põhimõttel, peab kaksikheeliks olema lahti keeratud ja algahelate vahel ei tohi olla vesiniksidemeid.

 Ainult sel juhul on DNA polümeraasid võimelised liikuma mööda lähteahelaid ja kasutama neid tütarahelate veatu sünteesi mallidena. Kuid paljudest miljonitest aluspaaridest koosnevate spiraalide täielik lahtikerimine on seotud nii märkimisväärse arvu pöörete ja selliste energiakuludega, mis rakutingimustes on võimatud. Seetõttu algab replikatsioon eukarüootides DNA molekuli mõnes kohas samaaegselt. Nimetatakse piirkonda kahe punkti vahel, kust algab tütarahelate süntees replikon. Ta on replikatsiooni ühik.
Ainult sel juhul on DNA polümeraasid võimelised liikuma mööda lähteahelaid ja kasutama neid tütarahelate veatu sünteesi mallidena. Kuid paljudest miljonitest aluspaaridest koosnevate spiraalide täielik lahtikerimine on seotud nii märkimisväärse arvu pöörete ja selliste energiakuludega, mis rakutingimustes on võimatud. Seetõttu algab replikatsioon eukarüootides DNA molekuli mõnes kohas samaaegselt. Nimetatakse piirkonda kahe punkti vahel, kust algab tütarahelate süntees replikon. Ta on replikatsiooni ühik.
Iga eukarüootse raku DNA molekul sisaldab palju replikone. Igas replikonis võib näha replikatsioonikahvlit – seda DNA molekuli osa, mis on spetsiaalsete ensüümide toimel juba lahti harutatud. Iga hargi ahel toimib täiendava tütarahela sünteesi mallina. Replikatsiooni ajal liigub kahvel mööda lähtemolekuli, samas kui uued DNA lõigud on lahti keeratud. Kuna DNA polümeraasid saavad maatriksi ahelaid mööda liikuda ainult ühes suunas ja ahelad on orienteeritud antiparalleelselt, sünteesitakse igas hargis korraga kaks erinevat ensümaatilist kompleksi. Veelgi enam, igas kahvlis kasvab üks tütarahel (juht) pidevalt ja teine (mahajäänud) ahel sünteesitakse mitme nukleotiidi pikkuste eraldi fragmentidega. Sellised ensüümid, mis said nime need avastanud Jaapani teadlase järgi Okazaki killud on seotud DNA ligaasiga, moodustades pideva ahela. DNA fragmentide tütarahelate moodustumise mehhanismi nimetatakse katkendlikuks.
5. Praimeri DNA polümeraasi vajadus ei suuda käivitada juhtiva ahela sünteesi ega ka mahajäänud ahela Okazaki fragmentide sünteesi. See saab üles ehitada ainult juba olemasoleva polünukleotiidahela, kinnitades desoksüribonukleotiide järjestikku oma 3'-OH otsa. Kust pärineb kasvava DNA ahela esialgne 5' ots? Seda sünteesib DNA matriitsil spetsiaalne RNA polümeraas, mida nimetatakse primaas(Inglise Primer - seeme). Ribonukleotiidpraimeri suurus on väike (alla 20 nukleotiidi) võrreldes DNA poimeraasi poolt moodustatud DNA ahela suurusega. Täitis oma Funktsioonid RNA praimer eemaldatakse spetsiaalse ensüümi abil ning selle käigus tekkinud tühimiku parandab DNA polümeraas, mis kasutab praimerina naabruses oleva Okazaki fragmendi 3'-OH otsa.
Lineaarsete DNA molekulide otste alareplikatsiooni probleem. Lineaarse alg-DNA molekuli mõlema ahela 3'-otsa komplementaarsete RNA praimerite eemaldamisel on tütarahelad lühemad kui 10–20 nukleotiidi. See on lineaarsete molekulide otste alareplikatsiooni probleem.
Lineaarsete DNA molekulide 3' otste alareplikatsiooni probleemi lahendavad eukarüootsed rakud spetsiaalse ensüümi abil - telomeraas.
Telomeraas on DNA polümeraas, mis täiendab kromosoomide 3'-terminaalseid DNA molekule lühikeste korduvate järjestustega. Need, paiknedes üksteise järel, moodustavad korrapärase kuni 10 tuhande nukleotiidi pikkuse terminaalse struktuuri. Telomeraas sisaldab lisaks valguosale RNA-d, mis toimib matriitsina DNA pikendamisel kordustega.
DNA molekulide otste pikenemise skeem. Esiteks toimub väljaulatuva DNA otsa komplementaarne seondumine telomeraasi RNA matriitsi saidiga, seejärel kogub telomeraas DNA, kasutades seemnena selle 3'-OH otsa ja matriitsina RNA-d, mis on osa ensüümist. Seda etappi nimetatakse pikenemiseks. Pärast seda toimub translokatsioon, s.o. DNA liikumine, mida pikendatakse ühe kordusega, ensüümi suhtes. Sellele järgneb pikenemine ja teine translokatsioon.
Selle tulemusena moodustuvad kromosoomide spetsiaalsed lõppstruktuurid. Need koosnevad korduvalt korduvatest lühikestest DNA järjestustest ja spetsiifilistest valkudest.
Lühikokkuvõte.
Eraldi tunnuse väljatöötamise võimaluse määramine
rakk või organism on GEEN.
Geenide ülekandmisel põlvkondade kaupa
tunnuste pärand vanematelt järglaste poolt.
GEENI kui funktsionaalse üksuse peamine OMADUS
pärilikkuse ja muutlikkuse materjal, on selle
keemiline organisatsioon.
Märkide moodustamisel on see nõutav
paljude ainete süntees
spetsiifiliste valkude järjestus
omadused.
Kinnisvara
valk
molekuli määrab aminohape
järjestus
teda
peptiid
ketid,
mis
antakse
DNA nukleotiidide järjestus.
Nukleiinhapped - DNA ja RNA
Nukleiinhapped DNA ja RNANukleiinhappeid on kahte tüüpi
Deoksüribonukleiinhapehape (DNA), koostises
mis sisaldab süsivesikuid desoksüriboosi
Ribonukleiinhape
hape (RNA)
mis sisaldab süsivesikute riboosi. DNA on raku suurim molekul.
See on palju suurem kui valgud ja RNA
Iga kromosoom = üks DNA molekul
23 inimese kromosoomi = 23 DNA molekuli
Pikim neist ≈ 8 cm
DNA on tekstimolekul. AT
selle nukleotiidjärjestused
salvestatakse kogu pärilik programm
organism
DNA asukoht rakus
TuumMitokondrid
plastiidid
Tuum
Kloroplast
Mitokondrid 1 DNA molekul
geen
teine geen
kromosoom
kromosoomid sisse
tuum
DNA
kamber
DNA funktsioonid
Säilitaminegeneetiline
teavet
Saade
geneetiline
teave alates
vanemad
järglased
Rakendamine
geneetiline
teave sisse
protsessi
elu andev
rakud ja
organism
Ankurdamine
Avastamise ajalugu
1. 1869 Friedrich Miescheravastas NK ja andis neile
nimi ("tuum" tuum).
2. 1905 Edwin Chargaff
uuritud nukleotiidi
NC koostis.
3. 1950 Rosalind
Franklin paigaldas,
kaheahelaline DNA.
Edwin
Chargaff
Rosalind
Franklin Röntgendifraktsioon
DNA portree -
kuulus foto 51
Rosalind Franklin
1920 - 1958http://www.bbc.co.uk/bbcfour/documentaries/features/rosalind- 1953
avatud
struktuur
DNA
kuupäev
sündi
molekulaarne
bioloogia
James
watson
Franciscus
karjuda James Dewey
Watson
Francis Harry Compton
Krikk
DNA molekule saab näha elektronmikroskoobiga
bakteriaalne plasmiidne DNA Reoviiruse DNAskaneeriv elekter. mikroskoop DNA isoleeritud
ühest kromosoomist
inimene
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/L/Laemmli
DNA on polümeermolekul, mis koosneb kahest komplementaarsest polünukleotiidahelast, mis on ühendatud vesiniksidemetega ja millel on pikk aeg
DNA molekuli struktuurDNA on polümeer
molekul, mis koosneb 2-st
tasuta
polünukleotiidahelad
ühendatud
vesinik
ühendused,
on suured ja
tohutu molekulaarne
mass.
DNA molekuli struktuur
Nukleotiidide aheladvormi
paremakäeline
10 paari mahulised spiraalid
alused igas pöördes
Ketid keerduvad ümber
üksteist ja ümber
ühine telg ja vorm
kaksikheeliks
Ketid on antiparalleelsed või
mitmesuunaline.
Järjekord
nukleotiidühendid
üks kett
vastupidine sellele sisse
teine
DNA skemaatiline struktuur
Nukleotiidid:1. Asuvad üksteisest eemal
sõber eemalt
0,34 nm
2. Mass ühe
nukleotiid on 345.
3. Spiraali laius 2nm
4. Need kogused
püsiv DNA struktuuri põhimõtted
5"
3"
AGA
G
G
T
C
AGA
AGA
C
5"
Ebakorrapärasus
T
C
C
AGA
kahekihiline
täiendavus
antiparallelism
G
T
T
G
3"
DNA struktuur
DNA on polümeer.Monomeerid on nukleotiidid.
Nukleotiid on keemiline ühend
kolme aine jäägid:
Nukleotiidi struktuur
lämmastikku sisaldav
põhjused:
- adeniin;
- guaniin;
- Tsütasiin
- Timin
Süsivesikud:
- desoksüriboos
Fosfori jäägid
happed (FA) Nukleotiid
5’
1’
fosfaat
lämmastikku sisaldav
alus -
üks 4-st
3’
Suhkur (riboos/desoksüriboos)
Lämmastikku sisaldavate aluste struktuuri skeemid.
DNA sisaldabjärgnev
lämmastikku sisaldav
põhjused:
Puriin
1. Adeniin,
2. Guaniin
pürimidiin
3. Timin
4. Tsütasiin
Sidemed sama DNA ahela nukleotiidide vahel
Rakendatudhariduse kaudu
fosforoeeter
vahelisi seoseid
desoksüriboos üks
nukleotiid ja jääk
fosforhappe
teine nukleotiid
Lingid ahelate vahel DNA molekulis
Rakendatudabiga
vesiniksidemed,
tekkimas
vahel
lämmastikku sisaldav
põhjustel
sissetulevad
sisse
ühend
erinevad ketid
1950. aasta Chargaffi reeglid
Erwin ChargaffChargaffi reeglite kohta andsid selgituse Watson ja Crick
DNA on ühendatud kahest ahelastpõhimõtte järgi
täiendavus
täiendavus
täiendavussee on
vastastikuse põhimõte
paaritud nukleotiidide sobitamine või võime
nukleotiidid paarituvad
Chargaffi reeglid
[ A ] + [ G ] \u003d [ T ] + [ C ] \u003d 50% Põhimõtetäiendavus:
AGA
-- -- --
T
G
-- -- ----
C
Tugevam
Nõrk
vesinik
ühendused!
Vastastikuse täiendavuse põhimõte
"replikatsiooni" omadus
DNA replikatsioon onkopeerimisprotsess
desoksüribonukleiinsed
hape, mis
toimub protsessi käigus
raku pooldumine.
Samas geneetiline
asjad krüpteeritud.
DNA-s kahekordistub ja
jagatud laste vahel
rakud.
DNA replikatsioon
sisseaega
replikatsioon
osa
molekulid
"ema" DNA rullub lahti kaheks ahelaks koos
kasutades spetsiaalset ensüümi ja seda
saavutatakse vaheliste vesiniksidemete katkestamisega
täiendavad lämmastiku alused:
adeniin-tüümiin ja guaniin-tsütosiin.
Edasi hajutatud ahelate igale nukleotiidile
DNA ensüüm DNA polümeraas kohandub
selle komplementaarne nukleotiid.
"replikatsiooni" omadus
Geneetiline kood
Pärilik teave salvestatakseNA molekulid järjestuse kujul
nukleotiidid.
Teatud
krundid
DNA ja RNA molekulid (viirustes ja faagides)
sisaldab teavet põhistruktuuri kohta
üks valk ja neid nimetatakse geenideks.
1 geen = 1 valgu molekul
Sellepärast
pärilik
teavet
mis
sisaldama
DNA
helistas
geneetiline.
Geneetilise koodi omadused:
Mitmekülgsusdiskreetsus
(kood
kolmikud
loetakse kogu RNA molekulist)
Spetsiifilisus (koodoni koodid ainult AK jaoks)
Koodi liiasus (mitu)
"reparatsiooni" vara
Remont – DNA molekuli võimeõige tekkimine selle ahelates
muudatusi.
DNA algse struktuuri taastamisel
Kaasatud on vähemalt 20 valku:
1. Tunneb ära muutunud DNA lõigud;
2. Eemaldage need ketist;
3. Taasta õige
nukleotiidjärjestus;
4. Õmble taastatud fragment
ülejäänud DNA molekul
Bibliograafia
Zahharov V.B. ja jne." Üldine bioloogia”Ruvinskiy A.O. Moskva "valgustus"
1993 “Üldbioloogia”
"Bioloogia tabelites ja diagrammides",
Bustard 2005
Internet: Google
Desoksüribonukleiinhape(DNA) on geneetilise teabe materiaalne kandja. See on kõrge molekulmassiga looduslik ühend, mis sisaldub elusorganismide rakkude tuumades. DNA molekulid koos histooni valkudega moodustavad aine kromosoomid. Histoonid on osa rakkude tuumadest ja osalevad rakutsükli erinevatel etappidel kromosoomide struktuuri säilitamises ja muutmises, geenide aktiivsuse reguleerimises. Teatud geenidele vastavad DNA molekulide eraldi lõigud. DNA molekul koosneb kahest polünukleotiidahelast, mis on keerdunud üksteise ümber spiraaliks (joonis 7.1). Ketid on ehitatud suurest hulgast nelja tüüpi monomeeridest - nukleotiidid mille spetsiifilisuse määrab üks neljast lämmastiku alusest: adeniin(AGA), tümiin(T), tsütosiin(C) ja guaniin(G). Kolme külgneva nukleotiidi kombinatsioon DNA ahela vormis geneetiline kood. Nukleotiidjärjestuse rikkumine DNA ahelas põhjustab kehas pärilikke muutusi - mutatsioonid. DNA reprodutseeritakse täpselt rakkude jagunemise käigus, mis tagab pärilike tunnuste ja spetsiifiliste ainevahetusvormide edasikandumise mitmes rakkude ja organismide põlvkondades.
Riis. 7.1. DNA molekuli struktuur.
DNA struktuurimudeli kaksikheeliksi kujul pakkusid 1953. aastal välja Ameerika biokeemik J. Watson (s. 1928) ja inglise biofüüsik ja geneetik F. Crick (s. 1916). Watson-Cricki mudel võimaldas selgitada paljusid DNA molekuli omadusi ja bioloogilisi funktsioone. Geneetilise koodi dešifreerimise eest pälvisid J. Watson, F. Crick ja inglise biofüüsik M. Wilkins (s. 1916), kes esmakordselt said kvaliteetse röntgenipildi DNA molekulist. Nobeli preemia 1962. aasta
DNA on hämmastav looduslik moodustis spiraalse sümmeetriaga. DNA ahela struktuuri pikad põimunud ahelad koosnevad suhkru- ja fosfaadimolekulidest. Lämmastikalused kinnituvad suhkrumolekulidele, moodustades ristsidemed kahe spiraalse ahela vahel. Piklik DNA molekul meenutab deformeerunud keerdtreppi. See on tõepoolest makromolekul: selle molekulmass võib ulatuda 10 9 . Vaatamata keerulisele struktuurile sisaldab DNA molekul ainult nelja lämmastiku alust: A, T, C, G. Adeniini ja tümiini vahel moodustuvad vesiniksidemed. Need sobivad struktuurilt nii kokku, et adeniin tunneb tümiini ära ja seondub sellega ning vastupidi. Tsütosiin ja guaniin on veel üks sarnast tüüpi paar. Nendes nukleotiidipaarides seostub sel viisil A alati T-ga ja C G-ga (joonis 7.2). See seos on kooskõlas täiendavuse põhimõtet. Aluspaaride arv: adeniin-tümiin ja tsütosiin-guaniin, näiteks inimestel, on suurejooneline: mõned teadlased usuvad, et neid on 3 miljardit, teised - rohkem kui 3,5 miljardit.

Lämmastikku sisaldavate aluste võime oma partnerit ära tunda toob kaasa suhkru-fosfaatahelate voltimise topeltheeliksi kujul, mille struktuur määratakse katseliselt röntgenvaatluste tulemusena. Lämmastikaluste vahelised vastasmõjud on väga spetsiifilised, seega saab spiraal tekkida ainult siis, kui mõlema ahela alusjärjestused on täiesti identsed.
Suhkrufosfaatrühm koos ühe lämmastiku alusega A, T, C või G, moodustades nukleotiid(Joonis 7.3) võib kujutada omamoodi ehitusplokina. Need plokid moodustavad DNA molekuli. Nukleotiidide järjestus kodeerib teavet DNA molekulis. See sisaldab teavet, mis on vajalik näiteks elusorganismile vajalike valkude tootmiseks.

DNA molekuli saab kopeerida ensüümide poolt katalüüsitava protsessi käigus replikatsioon, mis seisneb selle kahekordistamises. Replikatsioon lõhub vesiniksidemeid, moodustades üksikud ahelad, mis toimivad samade ehitusplokkide järjestuste ensümaatilise sünteesi mallina. Replikatsiooniprotsess hõlmab seega vanade katkemist ja uute vesiniksidemete moodustumist. Replikatsiooni alguses hakkavad kaks vastandlikku ahelat lahti kerima ja üksteisest eralduma (joonis 7.4). Lahtikeeramise hetkel seob ensüüm komplementaarsuse põhimõttel uued ahelad kahele vanale ahelale: uues ahelas on T vanas A vastas jne, mille tulemusena tekib kaks identset kaksikheeliksit. Selliste sidemete suhtelise hapruse tõttu toimub replikatsioon ilma suhkru-fosfaatahelate tugevamaid kovalentseid sidemeid purustamata. Geneetilise informatsiooni kodeerimine ja DNA molekuli replikatsioon on omavahel seotud peamised protsessid, mis on vajalikud elusorganismi arenguks.

Geneetiline teave on kodeeritud DNA nukleotiidide järjestusega. Põhitöö geneetilise koodi dešifreerimiseks tegid Ameerika biokeemikud M. Nirenberg (s. 1927), X. Koran (s. 1922) ja R. Holly (s. 1922); 1968. aasta Nobeli preemia laureaadid Kolm järjestikust nukleotiidi moodustavad geneetilise koodi ühiku, mida nimetatakse koodon. Iga koodon kodeerib kindlat aminohapet koguarv mis võrdub 20. DNA molekuli saab kujutada nukleotiiditähtede jadana, mis moodustavad teksti suurest hulgast neist, näiteks ACAT-TGGAG ... See tekst sisaldab teavet, mis määrab iga organismi eripära: inimene, delfiin jne. Kõigi elusolendite geneetiline kood, olgu see siis taim, loom või bakter, on sama. Näiteks koodon GGU kodeerib kõigis organismides aminohapet glütsiini. See geneetilise koodi tunnus koos kõigi valkude aminohappelise koostise sarnasusega annab tunnistust elu biokeemilisest ühtsusest, mis ilmselt peegeldab kõigi elusolendite päritolu ühest esivanemast.
1) i-RNA 2) t-RNA 3) DNA 4) kromosoom
A2. Inimese tütarrakkudes pärineb nahk nende paljunemise ajal emarakust:
täielik geneetiline teave
pool infost
veerand informatsioonist
õiget vastust pole
A3. DNA replikatsiooniga kaasneb keemiliste sidemete katkemine:
peptiid, aminohapete vahel
kovalentne, süsivesikute ja fosfaatide vahel
vesinik, lämmastikualuste vahel
iooniline, molekuli struktuuri sees
A4. DNA molekuli replikatsiooni käigus moodustub:
niit, mis on purunenud tütarmolekulide eraldi fragmentideks
molekul, mis koosneb kahest uuest DNA ahelast
molekul, millest pool koosneb mRNA ahelast
tütarmolekul, mis koosneb ühest vanast ja ühest uuest DNA ahelast
A5. Transkriptsioon on protsess:
1) DNA replikatsioon
2) i-RNA süntees
3) valgusüntees
4) t-RNA seondumine aminohappega
A6. Kui aminohapet kodeerib koodon UGG, siis DNA-s vastab see tripletile:
TCC 2) AGG 3) UCC 4) ACC
A7. Üks DNA kolmik sisaldab teavet:
Aminohappejärjestused valgu molekulis
Konkreetse AA asukoht valguahelas
Konkreetse organismi märk
Valguahelas sisalduvad aminohapped
A8. Translatsioonis osalevate tRNA-de arv on võrdne:
i-RNA koodonid, mis kodeerivad aminohappeid
Molekul i-RNA
Geenid, mis moodustavad DNA molekuli
Ribosoomidel sünteesitud valgud
A9. Raku eluperioodi jagunemisest jagunemiseni nimetatakse:
Interfaas 3) meioos
Mitoos 4) rakutsükkel
A10. Mitu kromatiidi sisaldub 8 kromosoomis, mis on nähtavad mitoosi metafaasis:
1) 6 2) 8 3) 12 4) 16
A11. Kromosoomide arv inimese somaatilistes rakkudes pärast mitoosi on:
1) 23 2) 46 3) 92 4) 44